Abstract
Heart failure results from the loss of structural integrity of the heart and/or a decrease in the rate of maximal ATP production. In cases of relatively preserved structural integrity, a decrease in ATP production in the mitochondria leads to a decrease in the cardiac stroke volume, thereby increasing the heart rate required to maintain the cardiac output. For many years, the exact location of this defect in the metabolic energy cycle remained elusive. Evidence is presented here to show that it is not a single metabolic substrate involved but rather the heavy isotope of hydrogen 2H, deuterium, that is jamming the ATP nanomotors slowing the rate of ATP production. During the digestion of a meal, the cardiac heart rate is shown to be very sensitive to the level of deuterium contained in the fatty acids recently consumed. During strenuous exercise in the fasting state, the enzyme adipose triglyceride lipase (ATGL) is found to mobilize the highest deuterium triglycerides more rapidly than the healthier lower deuterium triglycerides, converting the adipose tissue into a deuterium-depleted energy pool. This is believed to contribute to the low resting heart rates frequently observed in athletes. In vulnerable individuals, i.e., those weakened by disease(s) or space explorers in a weightless environment, the decreased ability to perform strenuous exercise leads to higher deuterium levels in their adipose tissue compromising their ATP production. In these individuals, maintaining healthy deuterium levels is best achieved by an increased intake of lower deuterium-containing foods.
Keywords
ATGL, adipose triglyceride lipase, ATP, ATPase nanomotor, cardiac, deuterium, deuterium-depletion, deuterium-depleted water, HFpEF, heart failure with preserved ejection fraction
Abbreviations
ATGL: adipose triglyceride lipase; CRP: C-reactive protein; D: deuterium; DDW: deuterium-depleted water; D/H: deuterium to hydrogen ratio; 1H: protium hydrogen isotope; 2H: deuterium hydrogen isotope; HFpEF: heart failure with preserved ejection fraction; HFrEF: heart failure with reduced ejection fraction; τ1/2: half-life; ECG: electrocardiogram; SLE: systemic lupus erythematosus; Pi: intracellular inorganic phosphate
Introduction
Heart failure is a very common and progressive disease diagnosed in roughly one percent of the population [1]. It results when the cardiac output is insufficient to meet the metabolic demands of the body [1, 2]. It can occur from the loss of structural integrity of the heart and/or a decrease in the rate of maximal ATP production. Detrimental changes to the cardiac structure occur in coronary artery disease, myocardial infarction, cardiomyopathy from substances/infections, chronic uncontrolled hypertension, chronic uncontrolled diabetes, cardiac birth defects, valvular damage, and high output demand from obesity [2]. In cases of relatively preserved structural integrity, the etiology has been attributed to a decrease in ATP production within the mitochondria from previous research studies [3]. These are frequently termed heart failure with reduced ejection fraction (HFrEF) when structural integrity is compromised and heart failure with preserved ejection fraction (HFpEF) when ATP production is slowed [4–8].
Any decrease in ATP production results in a decrease in the cardiac stroke volume, thereby increasing the heart rate required to maintain the cardiac output. When the conduction system is unable to pace the heart sufficiently enough to maintain the required cardiac output, HFpEF occurs. For many years the exact location of this defect within the metabolic energy pathway has remained elusive [4–7]. More recently, it was discovered that the heavy isotope of hydrogen 2H, deuterium, jams the ATP nanomotors at the end of the electron transport chain, decreasing the maximal rate of ATP production [9]. The level of deuterium found in food and water also varies and is systematically linked to the heart rate [3].
Deuterium 2H is a natural heavy isotope of hydrogen with an abundance of one 2H deuterium atom for every 6600 1H protium atoms on Earth. Tritium 3H is another heavy isotope of hydrogen but is found at extremely low levels in nature due to it being radioactive with a decay half-life τ1/2 of 12.32 years. The total concentration of deuterium in the human body is 12–14 mmol/L, which is higher than potassium, 3.5–5.1 mmol/L, indicating this isotope cannot be ignored in biochemical processes. When these deuterium isotopes interact with the ATP nanomotors in the mitochondria, cellular ATP production rates decrease [10–12]. The substitution of protium 1H with deuterium 2H in biochemical processes is called deuteration, and the reduction of the deuterium:protium (D/H) ratio below 1:6600 is called deuterium depletion or deupletion [13]. The discipline that studies and teaches how deuterium is discriminated and fractionated from protium in nature via biochemical reactions is called deutenomics [13]. Figure 1 illustrates the basic atomic structure of these hydrogen isotopes obtained from standard chemistry textbooks.
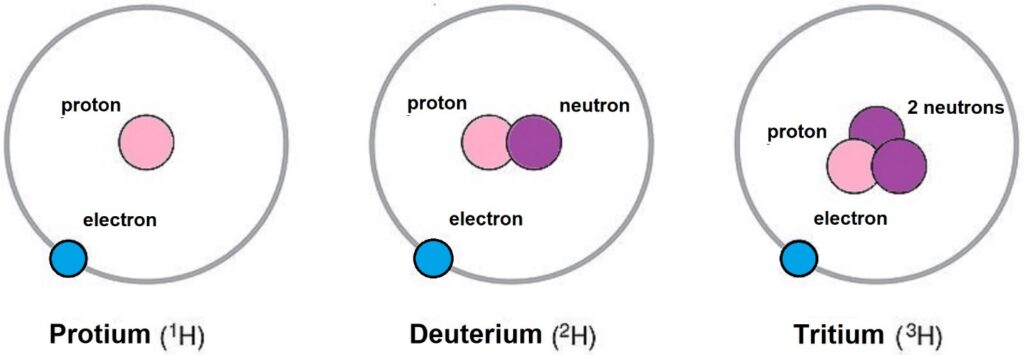 Figure 1: Isotopes of hydrogen. Protium is the most common isotope of hydrogen, with an abundance of 99.98% on Earth. Deuterium is a less common hydrogen isotope occurring with an abundance of 0.02% on Earth but still at levels 3x higher than potassium concentrations in the human body, 12–14 mmol/L. Tritium is a rare isotope due to being radioactive with a decay half-life, τ1/2, of 12.32 years. In nature, tritium is found mostly in the upper atmosphere, where cosmic rays interact with protium leaving two neutrons in the nucleus. The relative atomic masses are 1, 2, and 3, respectively.
Figure 1: Isotopes of hydrogen. Protium is the most common isotope of hydrogen, with an abundance of 99.98% on Earth. Deuterium is a less common hydrogen isotope occurring with an abundance of 0.02% on Earth but still at levels 3x higher than potassium concentrations in the human body, 12–14 mmol/L. Tritium is a rare isotope due to being radioactive with a decay half-life, τ1/2, of 12.32 years. In nature, tritium is found mostly in the upper atmosphere, where cosmic rays interact with protium leaving two neutrons in the nucleus. The relative atomic masses are 1, 2, and 3, respectively.
Deuterium 2H is not evenly distributed on the planet. This is primarily due to the freezing point of semi-heavy water HDO and heavy water D2O being 3.82°C rather than 0°C for H2O [14]. As a result, less deuterium reaches the colder polar regions of the planet compared to the equatorial regions. At the equator, the oceans have 155.76 ppm deuterium levels on average, which is known as the Vienna Standard Mean Ocean Water (VSMOW) [15]. At the south pole, this deuterium level falls to 85 ppm. Local climate also affects the deuterium levels, with the mountainous regions having lower deuterium levels than the deserts. Desert deuterium levels are further increased due to the vapor pressure of H2O being higher than HDO and/or D2O. For instance, the Sahara Desert is known to have deuterium levels approaching 180 ppm. Within the ocean ecosystem, deuterium levels in the top 500m of water near the shores are well documented to be lower near the ice caps indicating the ion concentrations of these regions are dictated by the freshwater runoff from the melting ice [16]. These are the regions where most cold-water seafood is caught.
The earliest known adverse effects of deuteration in living organisms were first reported in frogs and cows. In 1977, the sensitivity of frog retinas to light was found to decrease following exposure to deuterium [17]. By 1981, deuteration was found to decrease the ATP production in the mitochondria of cows [10–12]. This finding was instrumental in determining that a proton current along the electron-transport chain was directly involved in the production of ATP. However, the link between deuteration and human disease would take another three decades to develop.
In 1997, the Nobel Prize in chemistry was awarded to the three chemists Jens Christian Skou, John E. Walker, and Paul D. Boyer following their discovery that the ATP synthase was a rotary nanomotor spinning at approximately 9000 rpm [18]. By 2007, Olgun [9] calculated that the dissociation constant of deuterium from the Asp61 binding site of this ATP nanomotor was 35% that of protium, indicating the ATP nanomotor would significantly slow down lowering the overall ATP production rate as deuterium levels increase within the mitochondria intermembrane space.
More recently, the rates of remission from deuterium depletion have been studied in the lung [19], rare childhood [20], renal cell [21], and colorectal [22] cancers. Furthermore, the effect of deuterium levels on depression [23], diabetes mellitus [24, 25], and heart rate [3] has also been studied. Deuterium is also known to stabilize viral proteins and RNA to enhance their thermostability [26]. In this report, the effect of deuteration on the cardiac heart rate is presented. First, data are presented showing that deuterium depletion via a low carbohydrate (keto cycling diet) significantly decreases both the resting and exercise heart rates in a volunteer. Low heart rates have also been previously reported by others in a bodybuilder who adopted a ketogenic diet to lower his skeletal fat to 4.5% just prior to a contest [27]. Endurance athletes also develop low heart rates dropping as low as 27 bpm when their skeletal fat is decreased by frequent exercise [28]. Second, resting heart rates in six volunteers with normal heart function are presented showing that the resting heart rates systematically vary with the deuterium content from the last meal consumed. Finally, resting heart rates during the fasting state are presented showing the fasting heart rate is strongly influenced by the deuterium level stored in the adipose tissue. Data strongly suggest that exercise burns off the higher deuterium storage fats at a faster rate than the lower deuterium fats converting the adipose tissue into a deuterium-depleted energy pool.
A cardiac model explaining why the heart rate is sensitive to deuteration was previously reported in detail [3]. Summarized here, the volume of the heart’s left ventricular volume (V) is proportional to the circumference of the ventricle (c) to the third power, V = (4/3)πr3 = c3/(6π2), where r = average radius of the left ventricle. Scaling this equation to the average left ventricular volumes of males and females published in the literature [29] revealed that the heart rate would have to increase by 28% with a 5% drop in the maximal ATP production rate if no vascular compensation occurs [3]. This reveals why the resting heart rate can be utilized as a sensitive tool to detect the relative amounts of deuterium found in the fatty acids and proteins, the primary fuels utilized by the heart, consumed from the most recent meals. Figure 2 shows the compensated heart rates needed to maintain constant cardiac output for various decreases in the maximal ATP production rates. The ATP production is reduced by the deuterium isotopes reaching the ATP nanomotors.
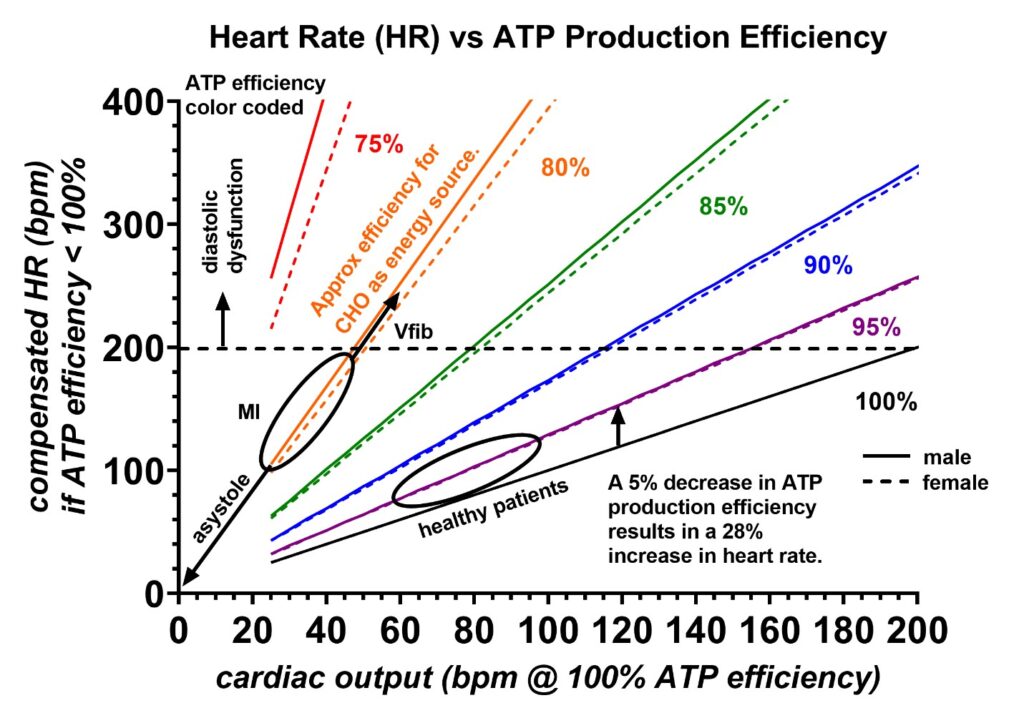 Figure 2: Compensated heart rates (vertical axis) needed to maintain various cardiac outputs (horizontal axis) are shown for different ATP production rate efficiencies (colors), assuming no compensation occurs in the peripheral vascular system. The cardiac outputs are in units of bpm at 100% ATP production efficiency. The vertical arrow indicates a 28% increase in heart rate with a 5% decrease in the ATP production rate. Carbohydrates generally contain more deuterium than fatty acids and are estimated to be 20% less efficient at producing ATP (orange). Compensated heart rates > 200 bpm are unsustainable for the dimensions of the human heart due to finite cardiac conduction velocities. This range in heart compensation will lead to diastolic dysfunction. During a myocardial infarction, the ATP production efficiency drops due to the cardiomyocytes being forced to utilize the inefficient anaerobic carbohydrate pathway. The higher deuterium levels in carbohydrates greatly increase the compensated heart rates, and if this becomes excessive, the conduction system cannot fully depolarize the ventricle per stroke, causing the ventricle to enter a state of contraction fasciculations known as ventricular fibrillation. Furthermore, if the ATP production drastically decreases into the inefficient lactate cycle, the heart can enter asystole. The region of normal cardiac output for a resting healthy individual is labeled. The prediction for males and females is also shown as solid and dashed lines, respectively. Finally, compensation in the peripheral vascular system tone will decrease the rate of SA node pacing required to maintain a constant cardiac output. Figure adopted from reference 3.
Figure 2: Compensated heart rates (vertical axis) needed to maintain various cardiac outputs (horizontal axis) are shown for different ATP production rate efficiencies (colors), assuming no compensation occurs in the peripheral vascular system. The cardiac outputs are in units of bpm at 100% ATP production efficiency. The vertical arrow indicates a 28% increase in heart rate with a 5% decrease in the ATP production rate. Carbohydrates generally contain more deuterium than fatty acids and are estimated to be 20% less efficient at producing ATP (orange). Compensated heart rates > 200 bpm are unsustainable for the dimensions of the human heart due to finite cardiac conduction velocities. This range in heart compensation will lead to diastolic dysfunction. During a myocardial infarction, the ATP production efficiency drops due to the cardiomyocytes being forced to utilize the inefficient anaerobic carbohydrate pathway. The higher deuterium levels in carbohydrates greatly increase the compensated heart rates, and if this becomes excessive, the conduction system cannot fully depolarize the ventricle per stroke, causing the ventricle to enter a state of contraction fasciculations known as ventricular fibrillation. Furthermore, if the ATP production drastically decreases into the inefficient lactate cycle, the heart can enter asystole. The region of normal cardiac output for a resting healthy individual is labeled. The prediction for males and females is also shown as solid and dashed lines, respectively. Finally, compensation in the peripheral vascular system tone will decrease the rate of SA node pacing required to maintain a constant cardiac output. Figure adopted from reference 3.
It is worth noting that decreased heart rates have also been associated with increased longevity [30], and this is likely due to lower deuterium levels in the fatty acids used to power all organs in addition to the heart. Lower deuterium levels reduce the need for the glycolysis lactate-producing metabolic pathway that only produces a net 2 ATP molecules compared to the 32 ATP molecules produced by the TCA cycle. Restated, up to 16x more efficient ATP production occurs in individuals with lower deuterium energy storage levels. Finally, careful in-home resting heart rate measurements offer a technique for monitoring potential toxic deuterium exposure from foods and/or drinking water.
Materials and Methods
A total of six volunteers, three males and three females, participated in these cardiac heart rate studies. They were selected on the basis of good cardiac health because a good understanding of the effect of deuterium on a healthy heart is essential for the full understanding of the effect of deuterium on a diseased heart. Furthermore, as of 2022, there was no known heart rate vs. deuterium data in the medical literature. None of the volunteers were on any cardiac medications at the time of the food tests, which would have complicated the data analysis. Their ECGs were all normal with no pathologic Q-waves, ST, or T-wave abnormalities, and none reported any angina. If admitted to a hospital, their GRACE scores would all fall between 34 and 98, indicating a low risk for cardiac mortality. Volunteers 1, 2, and 5 are medical field workers, volunteer 3 is an engineer, volunteer 4 is a physicist, and volunteer 6 is a weight trainer. Figure 3 shows a 12-lead ECG from volunteer 1, who provided much of the data published in this report.
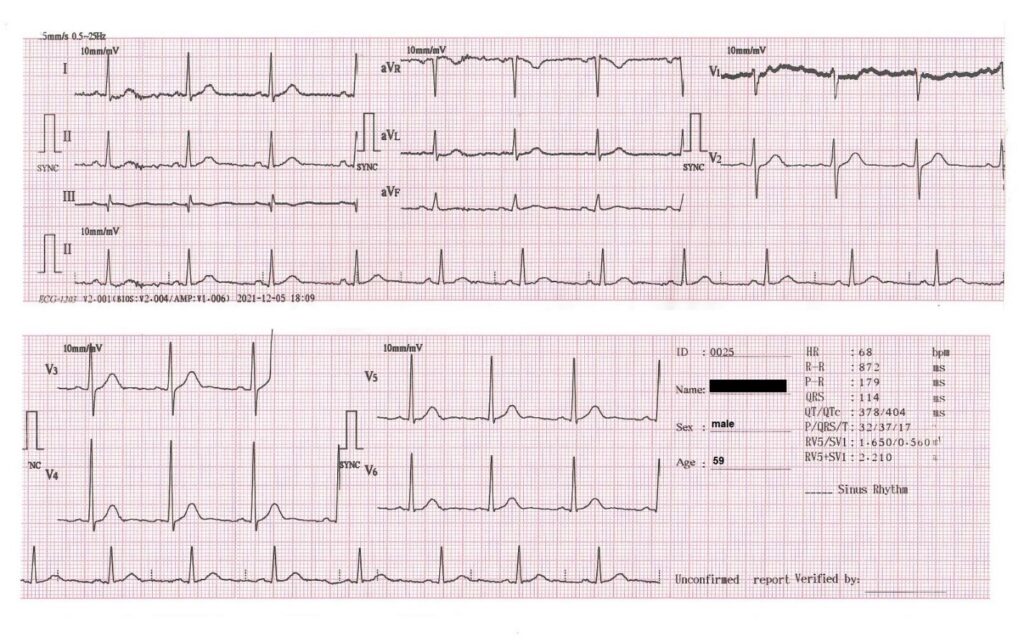 Figure 3: 12-lead electrocardiogram (ECG) from volunteer 1, who was a 59-year-old male at the time of this recording. The ECG shows normal sinus rhythm with no pathologic Q-waves, ST, or T-wave abnormalities. This recording was made 1 h after completing a 19.3 km hike.
Figure 3: 12-lead electrocardiogram (ECG) from volunteer 1, who was a 59-year-old male at the time of this recording. The ECG shows normal sinus rhythm with no pathologic Q-waves, ST, or T-wave abnormalities. This recording was made 1 h after completing a 19.3 km hike.
Volunteer 1 has maintained hiking log books dating back to the age of 18. These hiking logs have occasional peak heart rate entries beginning at the age of 24. These data were extremely valuable in producing some of the plots that appear later in this report. Furthermore, resting ECGs from previous physical exams remained available dating back to the age of 32. All these data allowed for the extraction of the historical resting and exercise heart rates over a period of four decades. It was the observation of a decrease in both the resting and exercise heart rates with the initiation of a keto cycling diet in this volunteer that prompted a detailed correlation of resting heart rates with the various meals consumed by the additional five volunteers.
All the volunteers recorded resting heart rates over a minimum period of two months immediately upon awakening from sleep or 3 h after dinner if they had a tendency to become hypoglycemic in the mornings. Volunteer 1 utilized an EMAY Model EMG-10 heart rhythm monitor to record 30-second rhythm strips indicating the 30-second averaged resting heart rates. Volunteers 2–6 used an over-the-counter pulse oximeter to record resting heart rates also averaged over 30 seconds. These were manually recorded. All the heart rate data from the six volunteers have been recorded for future use. In this report, volunteers 1 and 2 also recorded heart rate data for an additional period of 18 months beyond the first published report [3]. The technical specifications of the EMAY Model EMG-10 used by volunteer 1 are listed in Table 1.
| Lead | Standard 3 leads: Ⅰ Ⅱ Ⅲ |
| Calibration voltage | 1 mV ± 5% |
| Standard sensitivity | 10 mm/mV ± 5% |
| Amplitude frequency characteristic – standard | 10 Hz; 1 Hz~20 Hz; (+0.4 dB, -3 dB) |
| Noise level | ≤30 μV |
| Input impedance | ≥50 MΩ – CMRR: ≥60 dB |
| Scanning speed | 25 mm/s ± 5% |
| Sampling rate | 250 dots/s |
| HR measurement range | 30 bpm~300 bpm, error: ±1 bpm or 1% |
| Battery model | 602540P DC 3.7 V 530 mAh |
| Type of protection against electric shock | Internal power device |
| Metal electrode degree of protection against electric shock | Type BF applied part |
| Degree of waterproof | IP22 |
| Display | 1.77″ color TFT-LCD |
| Size | 100 mm (L) * 45 mm (W) * 15 mm (H) |
| Weight | about 60 g |
Table 1: EMAY model EMG-10 technical specifications.
Likewise, the technical specifications of the pulse oximeters used by volunteers 2–6 are given in Table 2. These pulse oximeters are FDA and CE-certified portable equipment featuring advanced technology for measuring and displaying functional oxygen saturation of arterial hemoglobin (%Sp02) and pulse rate (PR) obtained from the fingertip.
| Size | 58 mm x 36 mm x 33 mm, 2.3″ x 1.4″ x 1.3″ |
| Color | White/black and gray |
| Batteries | 2 x AAA |
| Resolution | ±1% |
| Measurement range | SpO2 (70–99%) and PR (30 bpm~240 bpm) |
| SpO2 accuracy | ±3% on the stage of 70% 99% |
| PR accuracy | ±1 bpm or ±1% |
Table 2: Finger pulse oximeter blood oxygen meter SpO2 heart rate monitor saturation.
Obtaining a typical heart rate data point is briefly described as follows. Each volunteer would select a specific food item to test and would consume that item at dinner time. The quantity needed was roughly 600 kcal or one-third of the basal metabolic rate per 24 h. The drawback is that the majority of calories need to be from a single food item, or the resulting heart rate data could be difficult to interpret. Therefore, in order to maintain a balanced diet, the other meals and snacks consumed earlier in the day would be selected to maintain a proper daily intake from each food group. The resting heart rates would then be measured after food is fully digested but before the body entered a fully fasting state. Heart rates in the fully fasting state reflect the deuterium level in the storage fats mobilized from the adipose tissue. For most individuals, the ideal measurement time is immediately before rising from bed. However, in volunteer 4, this time was selected as 3 h after dinner while this volunteer was studying for his graduate school classes. This was done because his high metabolism would put this volunteer into a hypoglycemic episode by the morning, increasing his heart rate from the adrenaline response. All data from the six volunteers were entered into spreadsheets which allowed for the production of the graphs presented in this report.
Cardiac outputs are well established to depend on multiple factors [31]. Until recently, this likely led many researchers to overlook the connection between the heart rate and deuterium levels. In these studies, to assure that none of the factors listed below were influencing the heart rate, a minimum of 5 resting heart rate readings on different dates (up to 25 in volunteer 1) were made for each food test. Consistency in the readings was typical for most tests except for an occasional upper respiratory infection that would increase the resting heart rates. These were not present during the first seven months of testing during the COVID-19 pandemic when other respiratory infections were low [32]. These data were recorded separately as an infection, and once the infection cleared, the resting heart rates would return to their original values, as indicated in Figure 4. The means and standard deviations in the resting heart rates for each food tested during the noninfectious periods are those that are included in all of the plots that follow. Table 3 lists the multiple factors that can influence the heart rate [33].
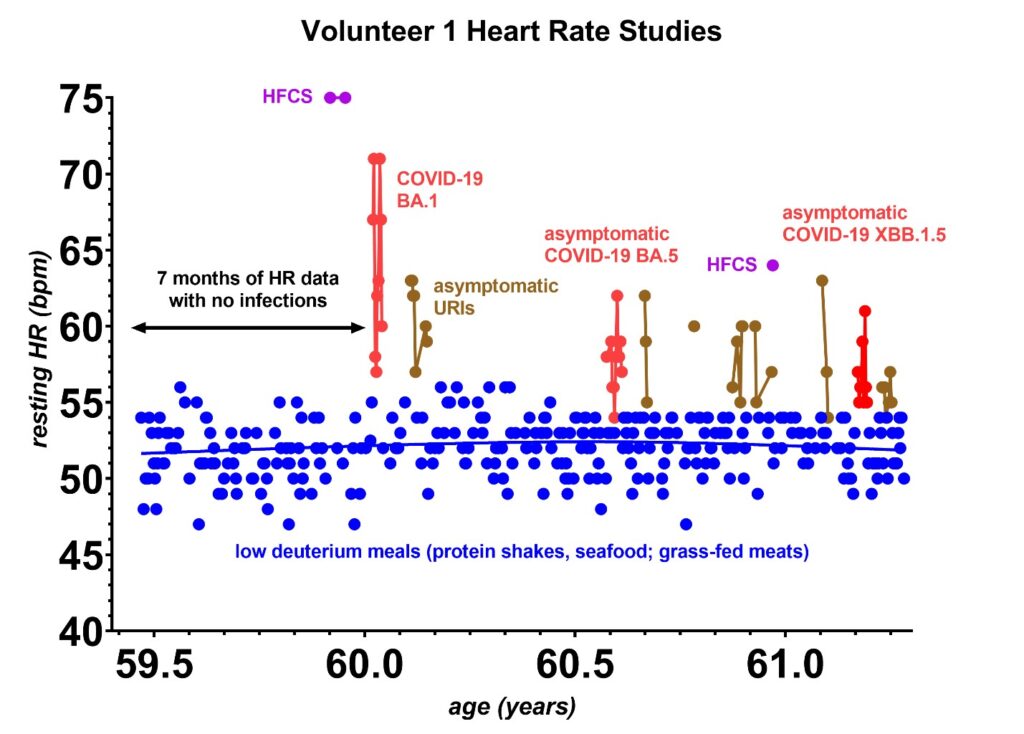 Figure 4: The first seven months of heart rate studies were made during the COVID-19 pandemic when circulating upper respiratory infections were low [32]. It was during these seven months that the heart rate patterns were established. Over the next year, upper respiratory infections, many of which were asymptomatic, began to progressively increase in prevalence. Most respiratory infections, although asymptomatic, do appear to increase the heart rate by ~5 bpm for 1–3 days (brown) and asymptomatic COVID-19 infections raise the heart rate by ~5 bpm for 9–10 days (red). At this time, one has to remain aware that such asymptomatic infections can appear at any time, dictating that each food tested needs to be retested on several occasions for consistency. The lower deuterium tests are marked in blue, and occasional high fructose corn syrup (HFCS) tests are marked as purple. The only symptomatic infection was the COVID-19 Omicron BA.1 at the age of 60.
Figure 4: The first seven months of heart rate studies were made during the COVID-19 pandemic when circulating upper respiratory infections were low [32]. It was during these seven months that the heart rate patterns were established. Over the next year, upper respiratory infections, many of which were asymptomatic, began to progressively increase in prevalence. Most respiratory infections, although asymptomatic, do appear to increase the heart rate by ~5 bpm for 1–3 days (brown) and asymptomatic COVID-19 infections raise the heart rate by ~5 bpm for 9–10 days (red). At this time, one has to remain aware that such asymptomatic infections can appear at any time, dictating that each food tested needs to be retested on several occasions for consistency. The lower deuterium tests are marked in blue, and occasional high fructose corn syrup (HFCS) tests are marked as purple. The only symptomatic infection was the COVID-19 Omicron BA.1 at the age of 60.
| 1. | Increased physical activity (exercise) |
| 2. | Medications/substances (sympathetic, parasympathetic, calcium channel blockers, alcohol, sedatives, opioids) |
| 3. | Infections including coronaviruses |
| 4. | Pain |
| 5. | Hypoglycemia |
| 6. | Tissue trauma |
| 7. | Blood volume depletion |
| 8. | Hyper/hypothermia |
| 9. | Smoking |
| 10. | Psychological (fear, anxiety, stress, anger) |
| 11. | Initial digestion of food |
| 12. | SA node and AV node disorders |
| 13. | Implanted cardiac pacemakers |
Table 3: Factors known to influence the heart rate [33].
A sample of four heart rate tests from volunteer 1 is shown in Figure 5. In these four tests, grass-fed bison, wild catch Argentine shrimp, grain-fed beef, and butter from grain-fed cows were recorded. More than 630 tests similar to these were recorded from volunteer 1. The range in heart rates recorded in these 630 tests varied from a minimum of 46 bpm to 75 bpm.
 Figure 5: Two-lead electrocardiogram (ECG) rhythms measured at lead position V4. The recordings were made with an EMAY Model EMG-10 just before rising from sleep. Resting heart rates shown here range from 46–61 bpm depending on the most recent food consumed during the previous evening. The top two ECGs compare a grass-fed bison ribeye to grain-fed beef, and the bottom two ECGs compare Argentine shrimp to grain-fed butter. Similar 30-second rhythm ECGs were recorded for each resting heart rate reported from volunteer 1 in this report.
Figure 5: Two-lead electrocardiogram (ECG) rhythms measured at lead position V4. The recordings were made with an EMAY Model EMG-10 just before rising from sleep. Resting heart rates shown here range from 46–61 bpm depending on the most recent food consumed during the previous evening. The top two ECGs compare a grass-fed bison ribeye to grain-fed beef, and the bottom two ECGs compare Argentine shrimp to grain-fed butter. Similar 30-second rhythm ECGs were recorded for each resting heart rate reported from volunteer 1 in this report.
Using mass spectrometry data already published [3, 34], the deuterium level calibration can be determined from a linear fit between the measured D/H ratios and the recorded resting heart rates from several food sources. The foods of known D/H (deuterium/hydrogen) ratios used in this report include: lard 116 ppm, casein protein shake 128 ppm, venison 128 ppm, grass-fed beef/elk fat 128 ppm, pea-based protein shake 129 ppm, extra virgin olive oil 130 ppm, free range chicken 130 ppm, grain-fed turkey 133 ppm, curd cottage cheese 136 ppm, grain-fed pork meat 148 ppm and high fructose corn syrup (HFCS)166 ppm. These were curve fit to each volunteer’s data to obtain the estimated fatty acid D/H level shown at the right axis of the graphs in the figures shown in this report. The specific curve fits were determined as follows and these are plotted in Figure 6:
Volunteer 1: D/H (ppm) = 1.5791 P + 48.852,
Volunteer 2: D/H (ppm) = 0.3773 P + 105.22,
Volunteer 3: D/H (ppm) = 1.1374 P + 60.28,
Volunteer 4: D/H (ppm) = 3.4786 P – 44.984,
Volunteer 5: D/H (ppm) = 2.2222 P – 12.00,
Volunteer 6: D/H (ppm) = 1.0456 P + 67.334, where P = resting heart rate.
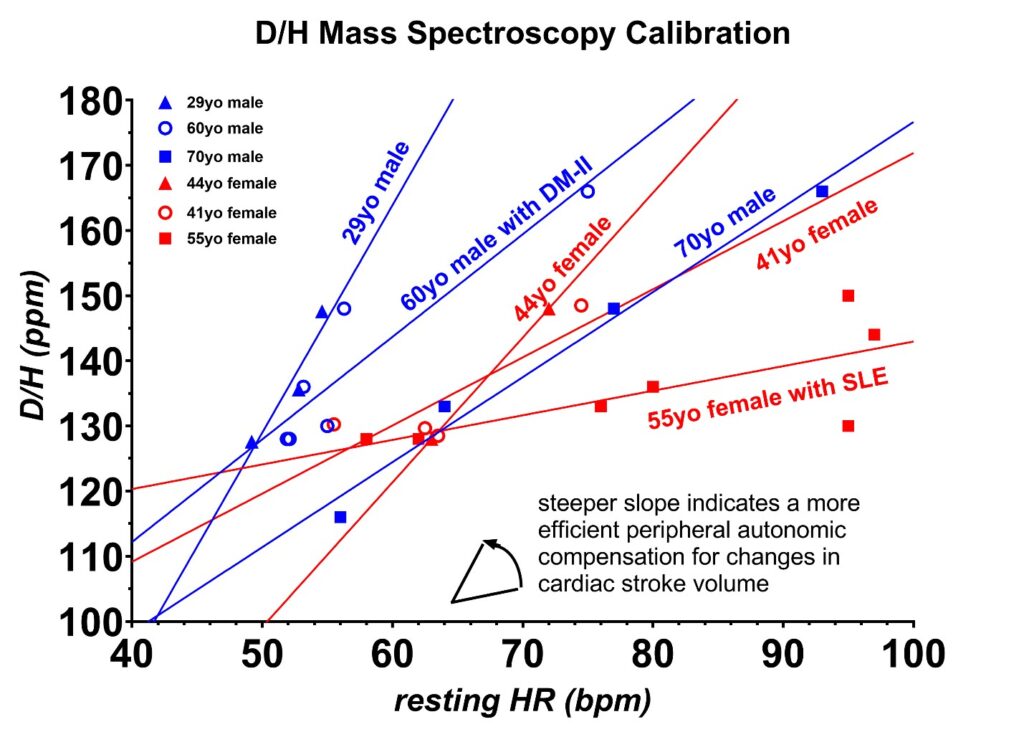 Figure 6: The deuterium calibration for the six volunteers is shown as a D/H ratio in parts per million (ppm) deuterium content vs. resting heart rates (HR) in beats per minute (bpm). The deuterium content from the foods in this plot was measured with mass spectroscopy, and these are listed in the text. The data shown in blue are from the three male volunteers, whereas the data shown in red are from the three female volunteers. All six volunteers had different calibration curves, which likely reflects the differences in the autonomic tone of the peripheral vascular system in each volunteer. The lowest heart rate range occurred in the youngest, healthiest volunteer, and the greatest heart rate range in the volunteer with a long history of systemic lupus erythematosus (SLE). The calibrations were repeated one year later in volunteers 1 and 2, and these had not shifted.
Figure 6: The deuterium calibration for the six volunteers is shown as a D/H ratio in parts per million (ppm) deuterium content vs. resting heart rates (HR) in beats per minute (bpm). The deuterium content from the foods in this plot was measured with mass spectroscopy, and these are listed in the text. The data shown in blue are from the three male volunteers, whereas the data shown in red are from the three female volunteers. All six volunteers had different calibration curves, which likely reflects the differences in the autonomic tone of the peripheral vascular system in each volunteer. The lowest heart rate range occurred in the youngest, healthiest volunteer, and the greatest heart rate range in the volunteer with a long history of systemic lupus erythematosus (SLE). The calibrations were repeated one year later in volunteers 1 and 2, and these had not shifted.
The differences in the slopes of the calibration curves are best explained as differences in the autonomic tone of the peripheral vascular system for each volunteer. The healthiest volunteer had the lowest change in resting heart rate with increasing deuterium while the volunteer who has a 30-year history of systemic lupus erythematosus (SLE) had the largest resting heart rate change with increasing deuterium. SLE is well known to adversely impact the peripheral vascular tone [35].
Results
Deuterium content of carbohydrates
Mass spectroscopy reveals that plants shift the higher concentrations of deuterium 2H preferentially towards the plant storage starches [34]. For instance, soluble corn fiber D/H levels have been reported at 155 ppm, wheat flour at 150 ppm, and potatoes at 143 ppm. Leafy vegetables are lower in deuterium, with an average leafy salad reported at 136 ppm D/H, cabbage at 142.5 ppm, and carrots at 142 ppm [34]. However, the heart does not directly run off carbohydrates but rather the fatty acids that are transported into the cardiomyocytes via the CD36 receptor and amino acids via the amino acid transporters [36]. Any effect of carbohydrate intake on the heart rate is an indirect consequence of the fats that have been produced from excessive carbohydrate intake modulated by elevated insulin levels. However, fructose is more rapidly converted to fatty acids and ceramides via first-pass hepatic metabolism as compared to glucose and these are found to increase the heart rate shortly after consumption [37]. Volunteer 1 discovered both a decrease in resting and peak exercise heart rates after adopting a cyclical ketogenic diet that decreased the overall amount of carbohydrates being consumed.
Figure 7 shows the physiologic changes that occurred in the resting heart rate (HR), systolic blood pressure (SBP), and diastolic blood pressure (DBP) of volunteer 1 between the age of 31 and 61 years. Data prior to age 53 were obtained from the available ECGs obtained from prior routine physicals made at rest in a fasting state. Data after age 53 were obtained from a portable EMAY Model EMG-10 ECG Monitor and a portable automated blood pressure monitor. The most significant change that is apparent is the dramatic decrease of 20 bpm in the resting heart rate when the keto cycling diet was initiated at age 53. The addition of regular consumption of deuterium-depleted water (DDW) starting four years later at age 57 decreased the resting heart rate by an additional 4 bpm. The weight of volunteer 1 is also given at several locations on the graph, and the deuterium level of the drinking water in ppm is also coded by the colors indicated. The blood pressure did vary slightly over the time period, but no systematic connection to the deuterium level was obvious. However, there was a marked improvement in the c-peptide at age 55 following the initiation of the keto cycling diet. There was also a gradual drop in the C-reactive protein (CRP) inflammatory marker from 2.5 mg/L at the time the keto cycling diet was first initiated to 0.36 mg/L four years after the DDW was added to the diet. These findings strongly suggest that the largest factor in deuterium depletion is achieved with a low carbohydrate diet but additional benefits do occur with the addition of DDW. The DDW does seem to lower the CRP which could be beneficial in patients at risk of coronary artery disease where elevated CRP levels are a risk factor for coronary artery blockages or autoimmune disorders [38].
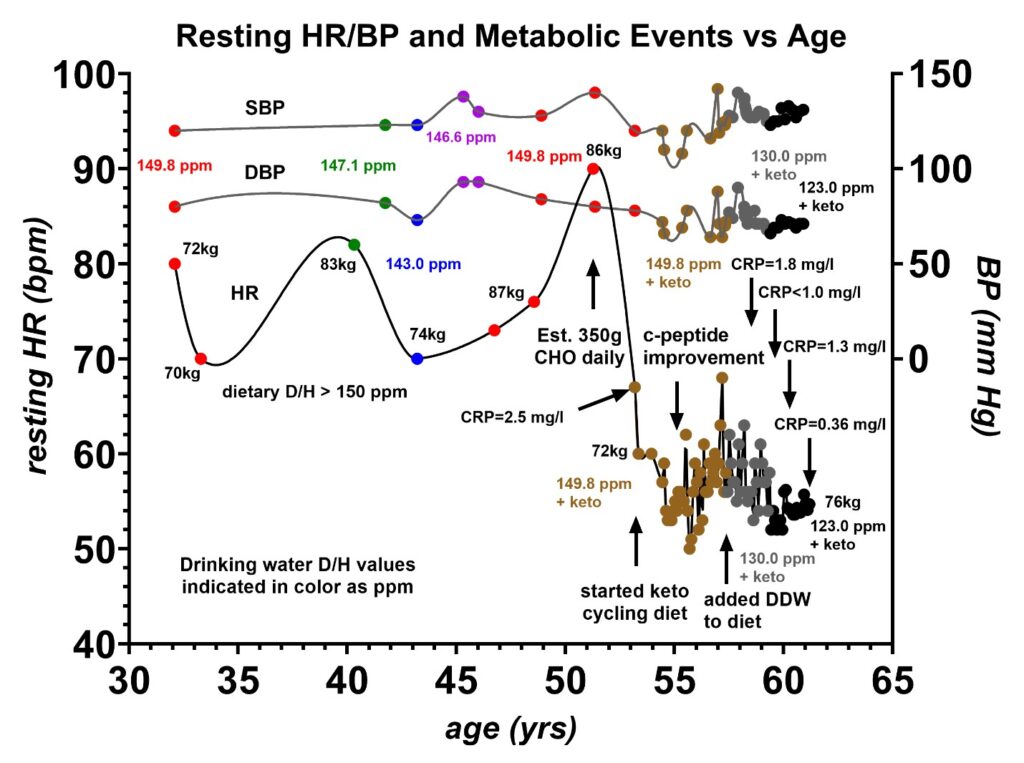 Figure 7: Resting heart rate (HR), systolic blood pressure (SBP), and diastolic blood pressure (DBP) vs. age. Volunteer 1 had a resting heart rate ranging between 70 and 90 beats per minute (bpm) prior to adopting a keto cycling diet. This heart rate dropped by 20 bpm to 50–68 bpm after the keto cycling diet was initiated. A 74% ejection fraction (EF) measured with nuclear SPECT at age 57 and later a 68% EF measured with an echocardiogram at age 61 indicate good contractility of the cardiac muscle. The addition of deuterium-depleted water (DDW) at age 57.5 years appears to have lowered the resting pulse by an additional 4 bpm indicating that the keto diet had the most influence on the resting heart rate. Prior to the keto diet, the predominant source of energy was from carbohydrates, where the deuterium content D/H was estimated to be greater than 150 ppm. The patient’s weight indicated at eight locations does not correlate with changes in the resting heart rate. The colors indicate the level of deuterium consumed in the drinking water as determined by mass spectroscopy. The C-reactive protein (CRP) inflammatory marker was measured at 2.5 mg/L when the keto cycling diet was first initiated at age 53, and this gradually dropped to very low levels, 0.36 mg/L, by the age of 61. A slight rebound to 1.3 mg/L briefly occurred following a COVID-19 infection at age 60.
Figure 7: Resting heart rate (HR), systolic blood pressure (SBP), and diastolic blood pressure (DBP) vs. age. Volunteer 1 had a resting heart rate ranging between 70 and 90 beats per minute (bpm) prior to adopting a keto cycling diet. This heart rate dropped by 20 bpm to 50–68 bpm after the keto cycling diet was initiated. A 74% ejection fraction (EF) measured with nuclear SPECT at age 57 and later a 68% EF measured with an echocardiogram at age 61 indicate good contractility of the cardiac muscle. The addition of deuterium-depleted water (DDW) at age 57.5 years appears to have lowered the resting pulse by an additional 4 bpm indicating that the keto diet had the most influence on the resting heart rate. Prior to the keto diet, the predominant source of energy was from carbohydrates, where the deuterium content D/H was estimated to be greater than 150 ppm. The patient’s weight indicated at eight locations does not correlate with changes in the resting heart rate. The colors indicate the level of deuterium consumed in the drinking water as determined by mass spectroscopy. The C-reactive protein (CRP) inflammatory marker was measured at 2.5 mg/L when the keto cycling diet was first initiated at age 53, and this gradually dropped to very low levels, 0.36 mg/L, by the age of 61. A slight rebound to 1.3 mg/L briefly occurred following a COVID-19 infection at age 60.
Figure 8 indicates the systematic changes in the maximum recorded heart rates during strenuous climbs between the age of 24 and 61. These exercise heart rates were obtained from volunteer 1’s hiking and climbing log books. The dates where type II diabetes was diagnosed, the keto cycling diet was initiated, c-peptide improvement occurred, and DDW was added to the diet are indicated [25]. Again, prior to age 53, the predominant energy source was carbohydrates (CHO), and after age 53, the predominant source was changed to fats via the keto cycling diet. Just above the x-axis, 13 rim-to-river backpacks in the Grand Canyon National Park are indicated to show that the drop in maximum heart rate was not due to a sudden improvement in physical fitness. Volunteer 1 has been in relatively good fitness his entire life. Both the resting and exercise heart rates appear to be strongly correlated to the primary energy source utilized, e.g., carbohydrates prior to age 53 and fats after age 53. Mass spectroscopy indicates that deuterium is more concentrated in the plant storage sugars, e.g., carbohydrates, as compared to the fats [34], suggesting that these drops in heart rates are due to systemic deuterium depletion in the skeletal fat.
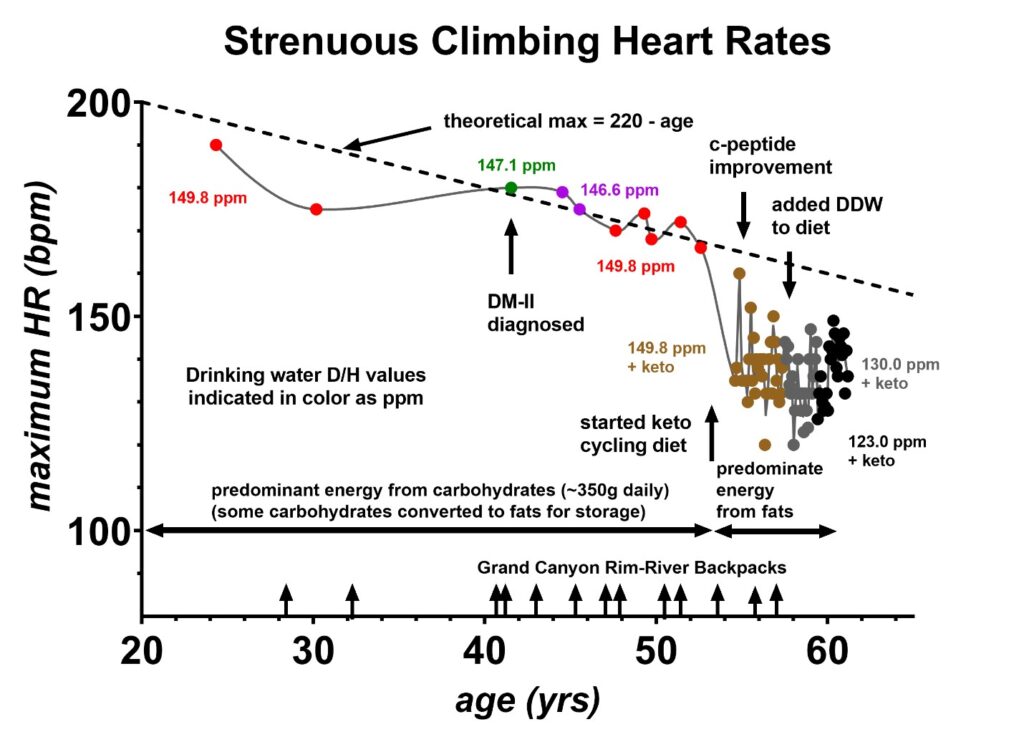 Figure 8: Peak exercise heart rates measured during strenuous climbs as a function of age. Volunteer 1 has hiking log books with peak heart rates dating back to age 24. Prior to age 53, the predominant energy source was carbohydrates, estimated at 350 g of daily intake. Following age 53, the predominant energy source was changed to fats at which time the peak exercise heart rates decreased below the theoretical maximum of (220-age). Between the age of 28 and 57 years, volunteer 1 made 13 backpacks in the Grand Canyon from the rim to the river and these trips are marked with the arrows just above the x-axis. These indicate that changes in overall physical fitness do not account for these maximum heart rate changes with age. Volunteer 1 was diagnosed with type II diabetes at age 41. Shortly after the keto cycling diet was initiated, a significant improvement in the c-peptide was observed at age 55, which was previously published [40].
Figure 8: Peak exercise heart rates measured during strenuous climbs as a function of age. Volunteer 1 has hiking log books with peak heart rates dating back to age 24. Prior to age 53, the predominant energy source was carbohydrates, estimated at 350 g of daily intake. Following age 53, the predominant energy source was changed to fats at which time the peak exercise heart rates decreased below the theoretical maximum of (220-age). Between the age of 28 and 57 years, volunteer 1 made 13 backpacks in the Grand Canyon from the rim to the river and these trips are marked with the arrows just above the x-axis. These indicate that changes in overall physical fitness do not account for these maximum heart rate changes with age. Volunteer 1 was diagnosed with type II diabetes at age 41. Shortly after the keto cycling diet was initiated, a significant improvement in the c-peptide was observed at age 55, which was previously published [40].
Deuterium content of fats and oils
Mass spectroscopy has also been used to measure the deuterium levels in various fats and oils. Many of these D/H values are published elsewhere and include curd cottage cheese at 136 ppm, extra virgin olive oil at 130 ppm, avocado oil at 130 ppm, grass-fed beef fat at 128 ppm, and grass-fed pork lard at 116 ppm [34]. Since the heart primarily utilizes fatty acids as an energy source [36], the greatest influence on the heart rate is expected to occur with the specific fat or oil recently consumed. As previously published, the heart rate was found to increase with higher deuterium levels due to the higher D/H fats decreasing the cardiac stroke volumes [3]. Resting heart rate data from various foods obtained by volunteer 1 are shown in Figures 9–14. Figure 9 compares the grass-fed land animals to grain-fed land animals, Figures 10 and 11 show the effect of various seafood and the latitude where these seafood are caught on the heart rate, Figures 12 and 13 compare the various nuts and seeds on the resting heart rate and Figure 14 compares various cooking oils on the resting heart rates. Where possible, the D/H value for cane sugar measured with mass spectroscopy was added for comparison. This is the value that one would expect for a resting heart rate if the cane sugar were to be converted to fat for utilization by the heart via the CD36 receptor [39].
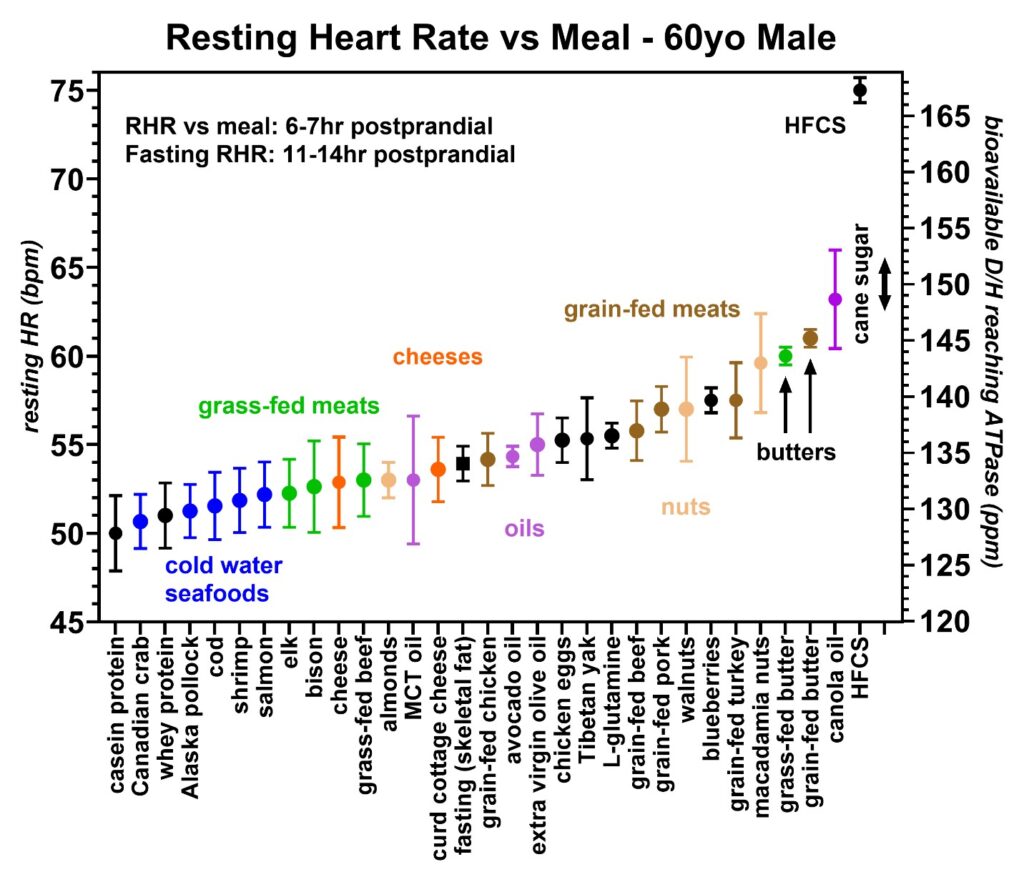 Figure 9: Resting heart rates recorded just prior to awakening that were collected over a 20-month period from volunteer 1 (59–61-year-old male). In each food group, the average and standard deviations are shown by circles and bars, respectively. A minimum of ten runs were made with each food group except for butter, canola oil, and high fructose corn syrup (HFCS), which were two runs each. The lowest heart rates were seen with the casein/whey protein shakes (black) followed by seafoods (blue), grass-fed meats (green), cheeses (orange), and grain-fed meats (brown). Some nuts and tree oils are also shown. The right axis is the estimated bioavailable D/H ratio as determined from the calibration equation. The black square near the center is the resting heart rate following fasting of 11–14 h which is believed to measure the D/H ratio in the skeletal fat. This fasting heart rate (HR) is very sensitive to average D/H intake and exercise level. The D/H range for cane sugar is also indicated and the measured D/H level of HFCS via first-pass hepatic metabolism is also shown. The important findings include that grass-fed animals give lower resting heart rates than grain-fed animals revealing that the relative D/H levels in these animals increase when these animals are fed grain products. Also, the cold water seafoods and protein shakes were among the lowest D/H foods recorded.
Figure 9: Resting heart rates recorded just prior to awakening that were collected over a 20-month period from volunteer 1 (59–61-year-old male). In each food group, the average and standard deviations are shown by circles and bars, respectively. A minimum of ten runs were made with each food group except for butter, canola oil, and high fructose corn syrup (HFCS), which were two runs each. The lowest heart rates were seen with the casein/whey protein shakes (black) followed by seafoods (blue), grass-fed meats (green), cheeses (orange), and grain-fed meats (brown). Some nuts and tree oils are also shown. The right axis is the estimated bioavailable D/H ratio as determined from the calibration equation. The black square near the center is the resting heart rate following fasting of 11–14 h which is believed to measure the D/H ratio in the skeletal fat. This fasting heart rate (HR) is very sensitive to average D/H intake and exercise level. The D/H range for cane sugar is also indicated and the measured D/H level of HFCS via first-pass hepatic metabolism is also shown. The important findings include that grass-fed animals give lower resting heart rates than grain-fed animals revealing that the relative D/H levels in these animals increase when these animals are fed grain products. Also, the cold water seafoods and protein shakes were among the lowest D/H foods recorded.
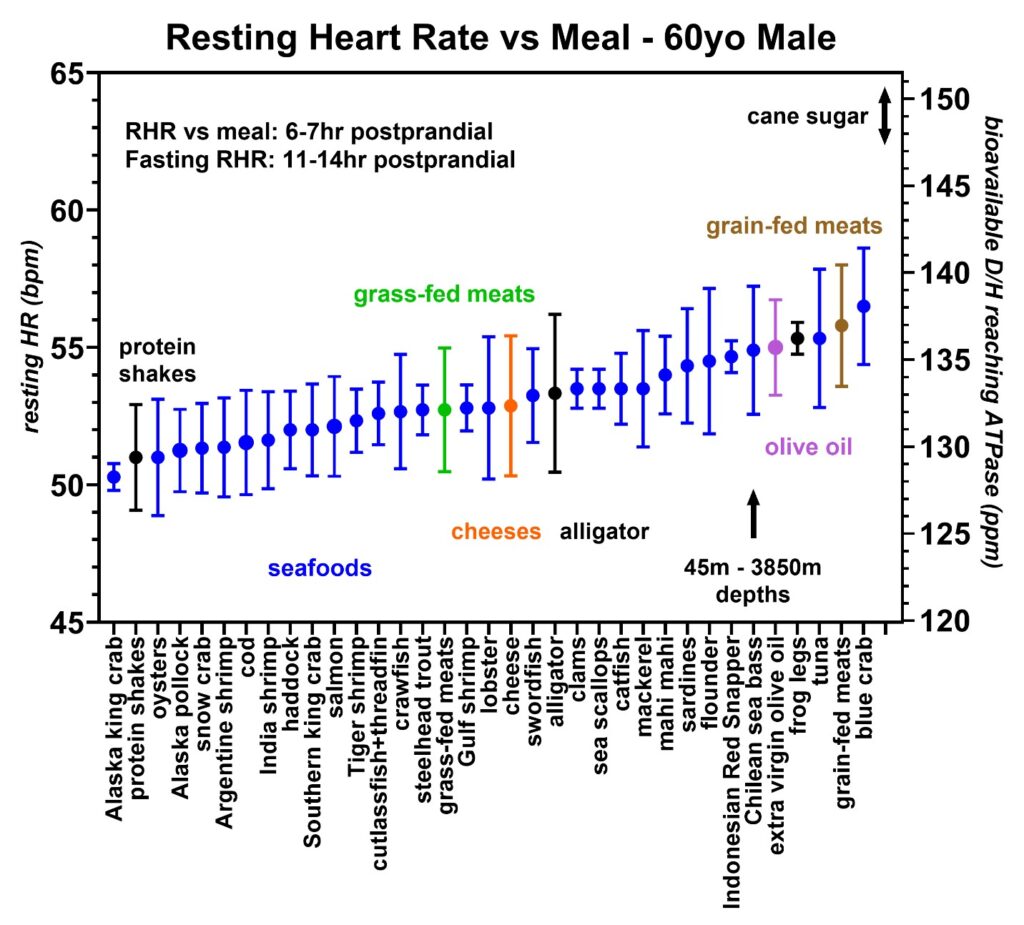 Figure 10: Resting heart rates recorded during the same 20-month period from volunteer 1 (59–61-year-old male) with an emphasis on the seafoods (blue). Protein shakes (black), grass-fed meats (green), cheeses (orange), alligator (black), olive oil (purple), and grain-fed meats (brown) are shown for comparison. Each species is found either at the surface of the water or shallow water (< 500 m) except for the Chilean sea bass which are caught in deeper waters ranging between 45–3850 m in depth. The D/H range of cane sugar is also indicated.
Figure 10: Resting heart rates recorded during the same 20-month period from volunteer 1 (59–61-year-old male) with an emphasis on the seafoods (blue). Protein shakes (black), grass-fed meats (green), cheeses (orange), alligator (black), olive oil (purple), and grain-fed meats (brown) are shown for comparison. Each species is found either at the surface of the water or shallow water (< 500 m) except for the Chilean sea bass which are caught in deeper waters ranging between 45–3850 m in depth. The D/H range of cane sugar is also indicated.
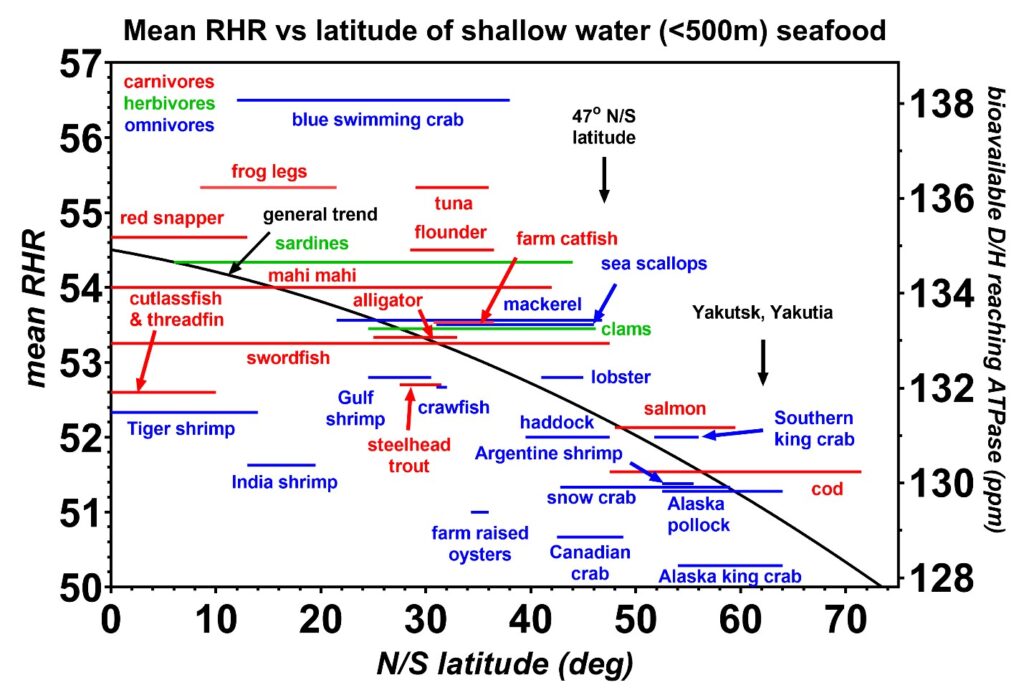 Figure 11: The mean resting heart rates (RHR) induced by the shallow seafoods (< 500 m depth) are plotted as a function of the range in latitude, either north or south of the equator, where each species is caught. All species are wild-caught except for the farm-raised catfish. Carnivores are indicated in red, herbivores in green, and omnivores in blue. The general trend shows that the colder water seafoods near the ice caps at latitudes above 47 degrees have the lowest D/H values. Small fish such as shrimp also have D/H values less than the trendline, suggesting these fish might spend a significant portion of their life cycle in the metabolic keto state which depletes their deuterium levels. One key finding implied from these data is that the lowest D/H values are obtained from fish closer to the melting ice where the measured ocean deuterium levels were previously reported by oceanographers to be lower [16]. It is also plausible that these lower D/H levels are strongly linked to the Ω3/Ω6 fatty acid ratios. Furthermore, the krill oils that are obtained from the colder Antarctic waters are likely to contain higher Ω3/Ω6 ratios than other varieties of fish oils. The latitude of Yakutia is also indicated where a high number of centurions are reported to live [41].
Figure 11: The mean resting heart rates (RHR) induced by the shallow seafoods (< 500 m depth) are plotted as a function of the range in latitude, either north or south of the equator, where each species is caught. All species are wild-caught except for the farm-raised catfish. Carnivores are indicated in red, herbivores in green, and omnivores in blue. The general trend shows that the colder water seafoods near the ice caps at latitudes above 47 degrees have the lowest D/H values. Small fish such as shrimp also have D/H values less than the trendline, suggesting these fish might spend a significant portion of their life cycle in the metabolic keto state which depletes their deuterium levels. One key finding implied from these data is that the lowest D/H values are obtained from fish closer to the melting ice where the measured ocean deuterium levels were previously reported by oceanographers to be lower [16]. It is also plausible that these lower D/H levels are strongly linked to the Ω3/Ω6 fatty acid ratios. Furthermore, the krill oils that are obtained from the colder Antarctic waters are likely to contain higher Ω3/Ω6 ratios than other varieties of fish oils. The latitude of Yakutia is also indicated where a high number of centurions are reported to live [41].
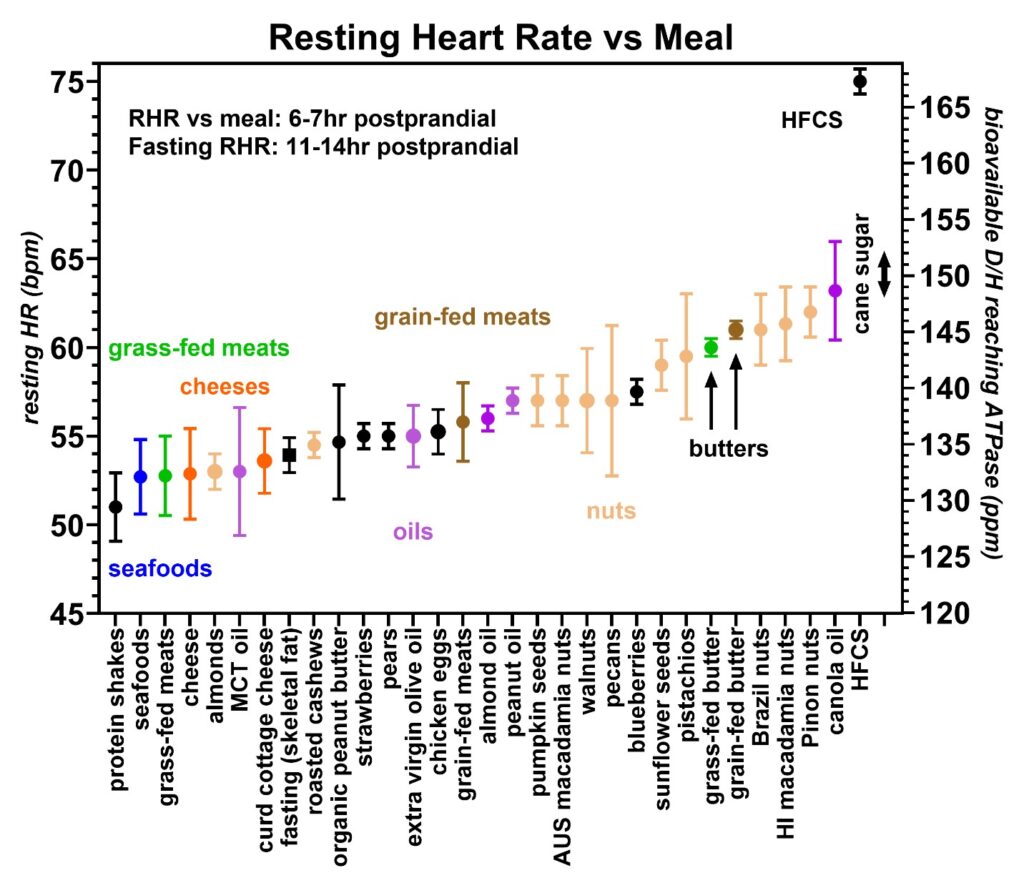 Figure 12: Resting heart rates recorded during the same 20-month period from volunteer 1 (59–61-year-old male) with an emphasis on the nuts and seeds (beige). The lowest D/H nuts and seeds include almonds and roasted cashews. Again, protein shakes (black), seafoods (blue), grass-fed meats (green), cheeses (orange), grain-fed meats (brown), and oils (purple) are indicated for comparison. Furthermore, high fructose corn syrup (HFCS) and cane sugar are indicated.
Figure 12: Resting heart rates recorded during the same 20-month period from volunteer 1 (59–61-year-old male) with an emphasis on the nuts and seeds (beige). The lowest D/H nuts and seeds include almonds and roasted cashews. Again, protein shakes (black), seafoods (blue), grass-fed meats (green), cheeses (orange), grain-fed meats (brown), and oils (purple) are indicated for comparison. Furthermore, high fructose corn syrup (HFCS) and cane sugar are indicated.
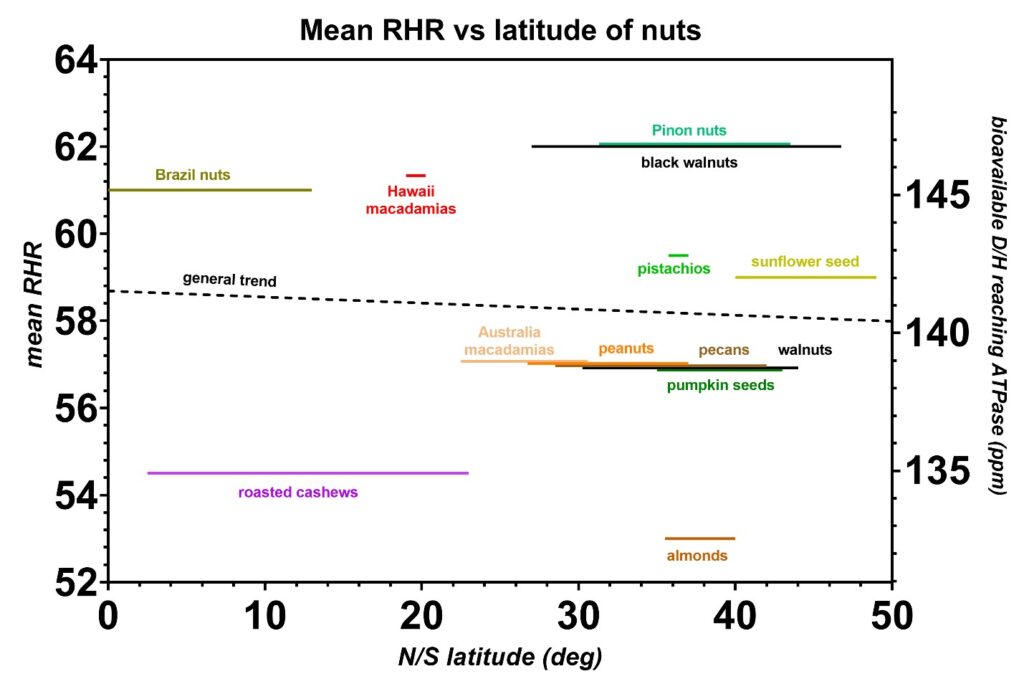 Figure 13: The mean resting heart rates (RHR) from the nuts and seeds plotted as a function of the range in latitude, either north or south of the equator, where each species is harvested. Unlike seafoods, the D/H levels in nuts and seeds are mostly independent of the latitude where they are harvested. The environmental D/H exposure likely depends mostly on the local climate and the relative location of the mountains to the harvesting sites. The general D/H trend in latitude is indicated (dashed line) and this trend is not statistically significant.
Figure 13: The mean resting heart rates (RHR) from the nuts and seeds plotted as a function of the range in latitude, either north or south of the equator, where each species is harvested. Unlike seafoods, the D/H levels in nuts and seeds are mostly independent of the latitude where they are harvested. The environmental D/H exposure likely depends mostly on the local climate and the relative location of the mountains to the harvesting sites. The general D/H trend in latitude is indicated (dashed line) and this trend is not statistically significant.
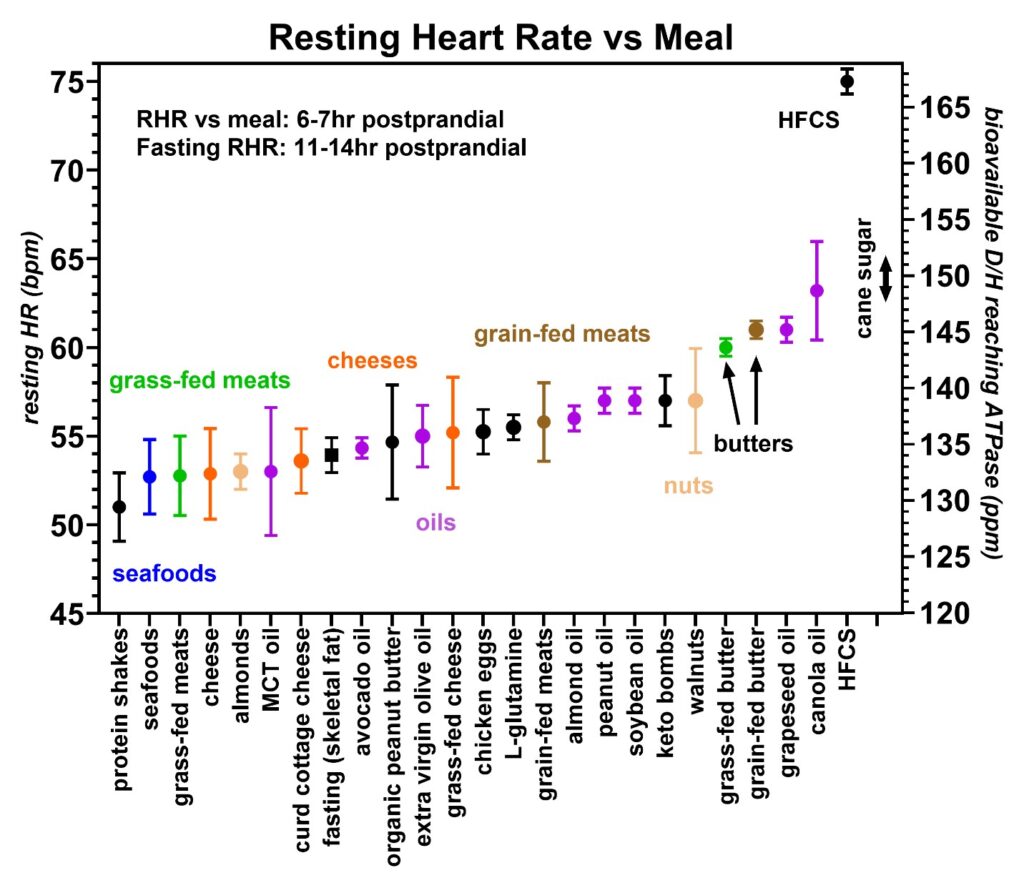 Figure 14: Resting heart rates recorded during the same 20-month period from volunteer 1 (59–61-year-old male) with an emphasis on the cooking oils (purple). The healthiest oils are MCT (refined 6 and 8-carbon oils from coconut oil), extra virgin olive oil, and avocado oil. Protein shakes (black), grass-fed meats (green), cheeses (orange), grain-fed meats (brown), and nuts (beige) are also included for comparison. High fructose corn syrup (HFCS) and cane sugar are again indicated.
Figure 14: Resting heart rates recorded during the same 20-month period from volunteer 1 (59–61-year-old male) with an emphasis on the cooking oils (purple). The healthiest oils are MCT (refined 6 and 8-carbon oils from coconut oil), extra virgin olive oil, and avocado oil. Protein shakes (black), grass-fed meats (green), cheeses (orange), grain-fed meats (brown), and nuts (beige) are also included for comparison. High fructose corn syrup (HFCS) and cane sugar are again indicated.
The resting heart rate to D/H food patterns presented in the next six figures was obtained from volunteers 2–6. These were obtained to see if the same patterns found in volunteer 1 occurred in a wider population. Figures 15 and 16 were collected from volunteer 2 who is a 55-year-old female. Figure 17 was collected from a 70-year-old male, Figure 18 from a 29-year-old male, Figure 19 from a 44-year-old female, and Figure 20 from a 41-year-old female. In all the volunteers, the patterns were consistent with volunteer 1, with the protein shakes giving the lowest resting heart rates, followed by grass-fed meats and then grain-fed meats. In volunteer 2, the resting heart rates induced by the colder water seafood were also found to be lower than the warmer water seafood with the same pattern as found earlier in volunteer 1; however, these seafood data from volunteer 2 were more limited due to an allergy to certain shellfish. However, the rates of increase in the resting heart rates with increasing D/H values were found to vary between individuals and most likely indicate differences in the autonomic tone of the peripheral vascular system in each volunteer, with the youngest, healthiest volunteer (Figure 18) showing the lowest rate of change and volunteer 2 with a history of SLE with the highest rate of change [3]. Again, where possible, the D/H value for cane sugar measured with mass spectroscopy was added for comparison.
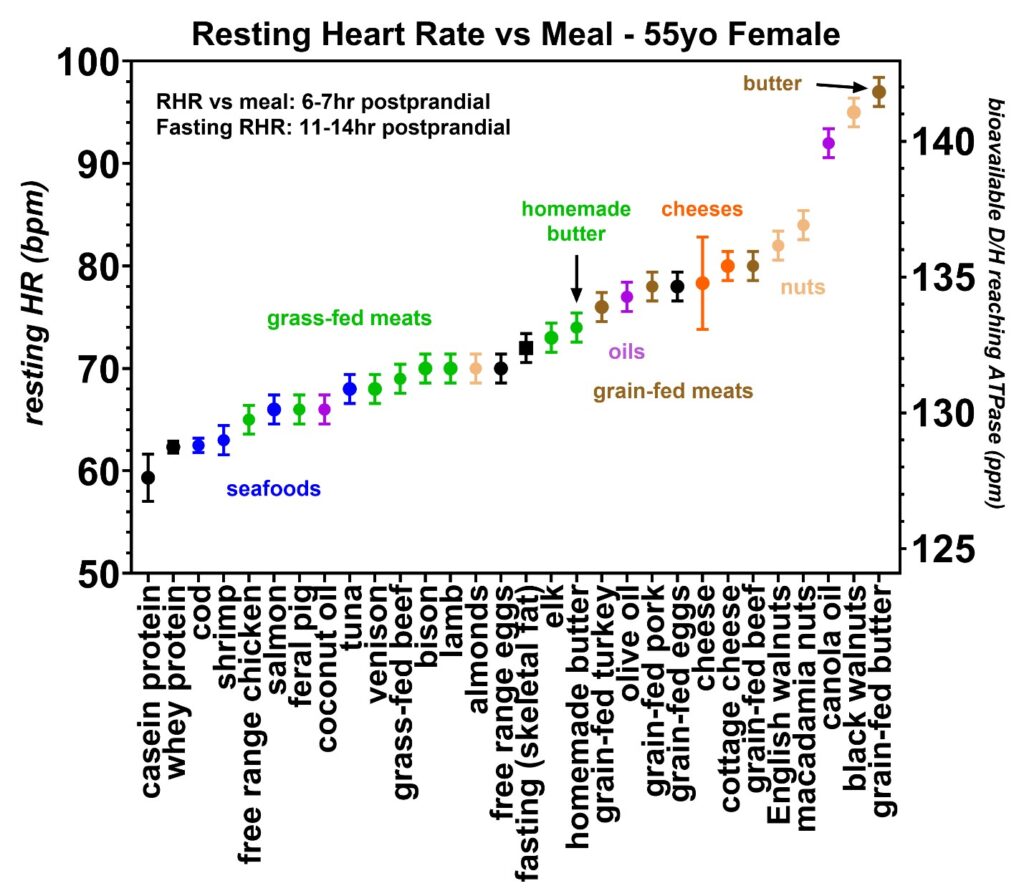 Figure 15: Resting heart rates recorded just prior to awakening obtained over a 20-month period from volunteer 2 (55–56-year-old female). In each food group, the average and standard deviations are also shown by circles and bars, respectively. Similar to volunteer 1, the lowest resting heart rates occurred with the protein shakes (black) followed by seafoods (blue), grass or naturally-fed meats (green), grain-fed meats (brown), cheeses (orange), and nuts (beige). Volunteer 2 also lived in a farming community and had access to free-range chicken meat, feral pigs, venison, and homemade butter. These four categories all fall into the grass- or natural-fed meat range of resting heart rates, indicating lower deuterium levels than the commercially available meats. This volunteer had the greatest range in resting heart rates which is believed to be due to her long history of systemic lupus erythematosus (SLE), as discussed in the report. Volunteer 2 also showed lower heart rates with the homemade butter made with natural ingredients compared to the commercially available butter. Although unclear, the commercially available butter may have a certain degree of hydrogenation to improve the shelf life, and hydrogenation likely utilizes commercially available hydrogen with a D/H ~ 155.76 ppm (equatorial seawater) [15].
Figure 15: Resting heart rates recorded just prior to awakening obtained over a 20-month period from volunteer 2 (55–56-year-old female). In each food group, the average and standard deviations are also shown by circles and bars, respectively. Similar to volunteer 1, the lowest resting heart rates occurred with the protein shakes (black) followed by seafoods (blue), grass or naturally-fed meats (green), grain-fed meats (brown), cheeses (orange), and nuts (beige). Volunteer 2 also lived in a farming community and had access to free-range chicken meat, feral pigs, venison, and homemade butter. These four categories all fall into the grass- or natural-fed meat range of resting heart rates, indicating lower deuterium levels than the commercially available meats. This volunteer had the greatest range in resting heart rates which is believed to be due to her long history of systemic lupus erythematosus (SLE), as discussed in the report. Volunteer 2 also showed lower heart rates with the homemade butter made with natural ingredients compared to the commercially available butter. Although unclear, the commercially available butter may have a certain degree of hydrogenation to improve the shelf life, and hydrogenation likely utilizes commercially available hydrogen with a D/H ~ 155.76 ppm (equatorial seawater) [15].
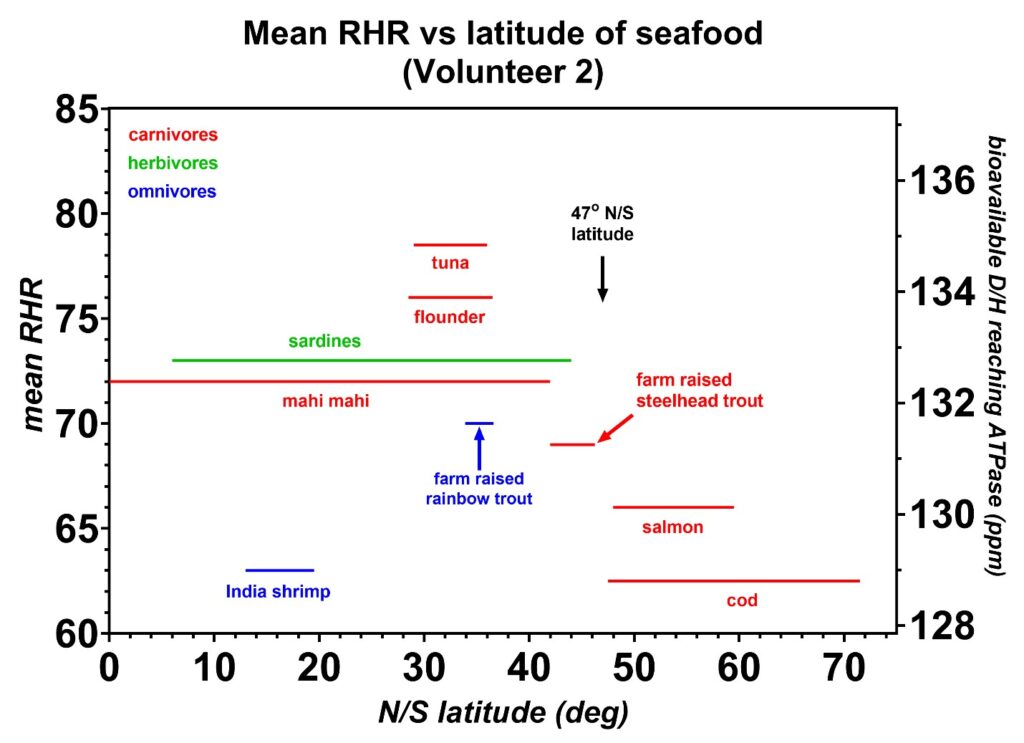 Figure 16: The mean resting heart rates (RHR) induced by the shallow seafoods (< 500 m depth) are plotted as a function of the range in latitude, either north or south of the equator, where each species is caught. All species are wild-caught except for the farm-raised steelhead trout and rainbow trout. Carnivores are indicated in red, herbivores in green, and omnivores in blue. Similar to volunteer 1 (Figure 11), the colder water seafoods near the ice caps at latitudes above 47 degrees have the lowest D/H values.
Figure 16: The mean resting heart rates (RHR) induced by the shallow seafoods (< 500 m depth) are plotted as a function of the range in latitude, either north or south of the equator, where each species is caught. All species are wild-caught except for the farm-raised steelhead trout and rainbow trout. Carnivores are indicated in red, herbivores in green, and omnivores in blue. Similar to volunteer 1 (Figure 11), the colder water seafoods near the ice caps at latitudes above 47 degrees have the lowest D/H values.
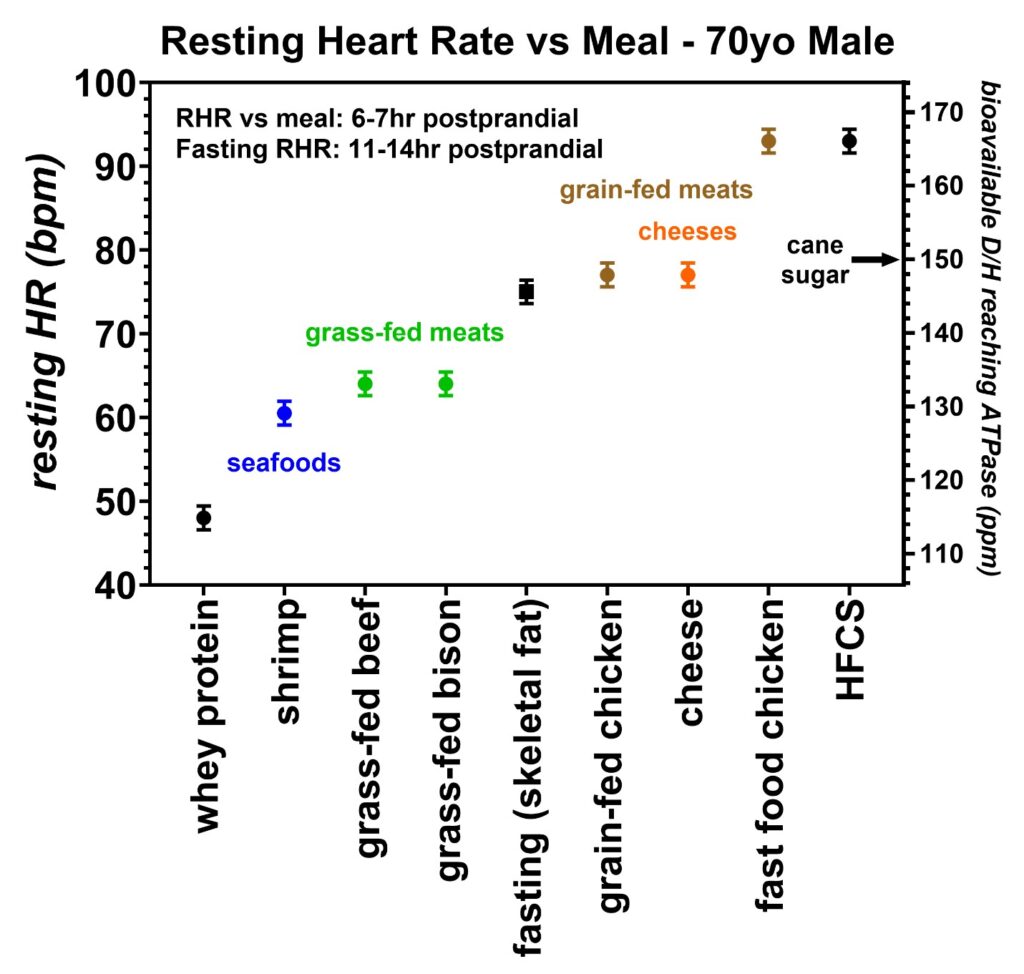 Figure 17: Resting heart rates recorded just prior to awakening over a five-month period collected by volunteer 3 (70-year-old male). The trend in resting heart rates was lowest in whey proteins (black), followed by shrimp (blue), grass-fed meats (green), grain-fed chickens (brown), cheeses (orange), and high fructose corn syrup (HFCS) (black). The estimated bioavailable D/H ratio is also given on the right y-axis as determined by the calibration equation. One of the fast-food chickens tested induced an elevated resting heart rate suggesting this food may have been prepared with either a high-deuterium cooking oil or HFCS. The D/H ratio for cane sugar measured with mass spectroscopy is also indicated.
Figure 17: Resting heart rates recorded just prior to awakening over a five-month period collected by volunteer 3 (70-year-old male). The trend in resting heart rates was lowest in whey proteins (black), followed by shrimp (blue), grass-fed meats (green), grain-fed chickens (brown), cheeses (orange), and high fructose corn syrup (HFCS) (black). The estimated bioavailable D/H ratio is also given on the right y-axis as determined by the calibration equation. One of the fast-food chickens tested induced an elevated resting heart rate suggesting this food may have been prepared with either a high-deuterium cooking oil or HFCS. The D/H ratio for cane sugar measured with mass spectroscopy is also indicated.
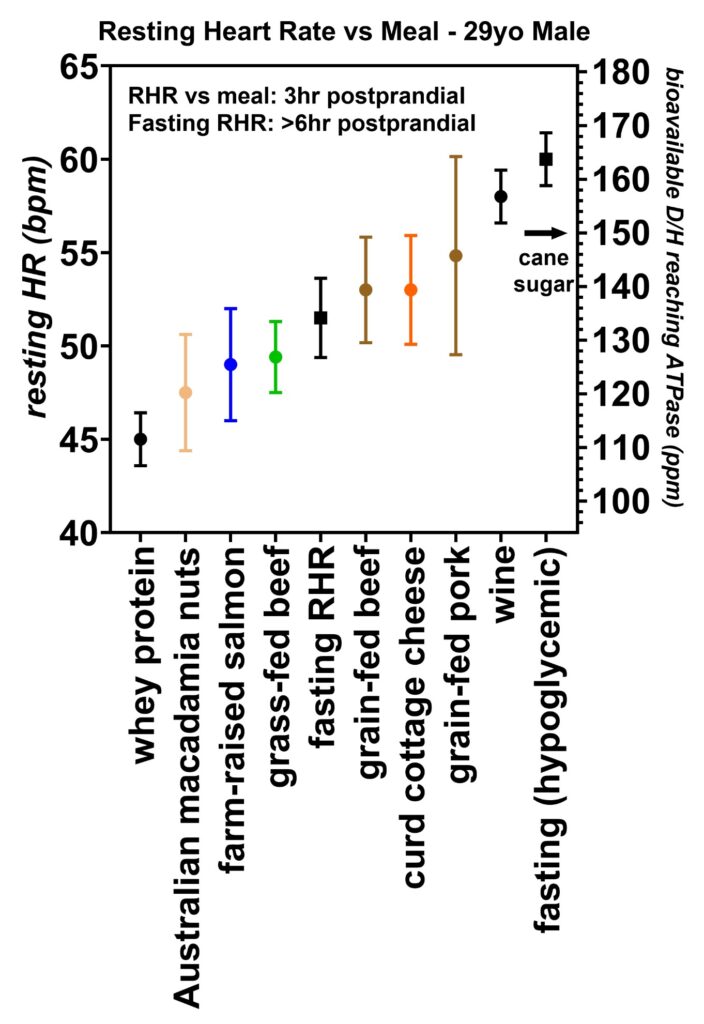 Figure 18: Resting heart rates recorded 3 h postprandial over a three-month period from volunteer 4 (29-year-old male). This volunteer was the youngest and most athletic of the six volunteers tested. His metabolism was sufficiently high enough that he became mildly hypoglycemic in the mornings prompting him to collect the resting heart rate data at 3 h postprandial. As with the other volunteers, the whey proteins led to the lowest resting heart rates, followed by salmon, grass-fed beef, and then grain-fed pork. A test of Australian macadamia nuts did reveal an unexpectedly low heart rate, and he also reported chills after consuming these nuts. In regards to the low resting heart rate, it is unclear if these nuts are lower in deuterium or if some type of allergic reaction led to this low heart rate. This volunteer also tested some wine and found an increased heart rate that corresponds to a D/H ~ 156 ppm which is the same as the mean equatorial seawater [16]. His typical fasting hypoglycemic HR values are shown to the right of the figure. The D/H ratio for cane sugar measured with mass spectroscopy is also indicated.
Figure 18: Resting heart rates recorded 3 h postprandial over a three-month period from volunteer 4 (29-year-old male). This volunteer was the youngest and most athletic of the six volunteers tested. His metabolism was sufficiently high enough that he became mildly hypoglycemic in the mornings prompting him to collect the resting heart rate data at 3 h postprandial. As with the other volunteers, the whey proteins led to the lowest resting heart rates, followed by salmon, grass-fed beef, and then grain-fed pork. A test of Australian macadamia nuts did reveal an unexpectedly low heart rate, and he also reported chills after consuming these nuts. In regards to the low resting heart rate, it is unclear if these nuts are lower in deuterium or if some type of allergic reaction led to this low heart rate. This volunteer also tested some wine and found an increased heart rate that corresponds to a D/H ~ 156 ppm which is the same as the mean equatorial seawater [16]. His typical fasting hypoglycemic HR values are shown to the right of the figure. The D/H ratio for cane sugar measured with mass spectroscopy is also indicated.
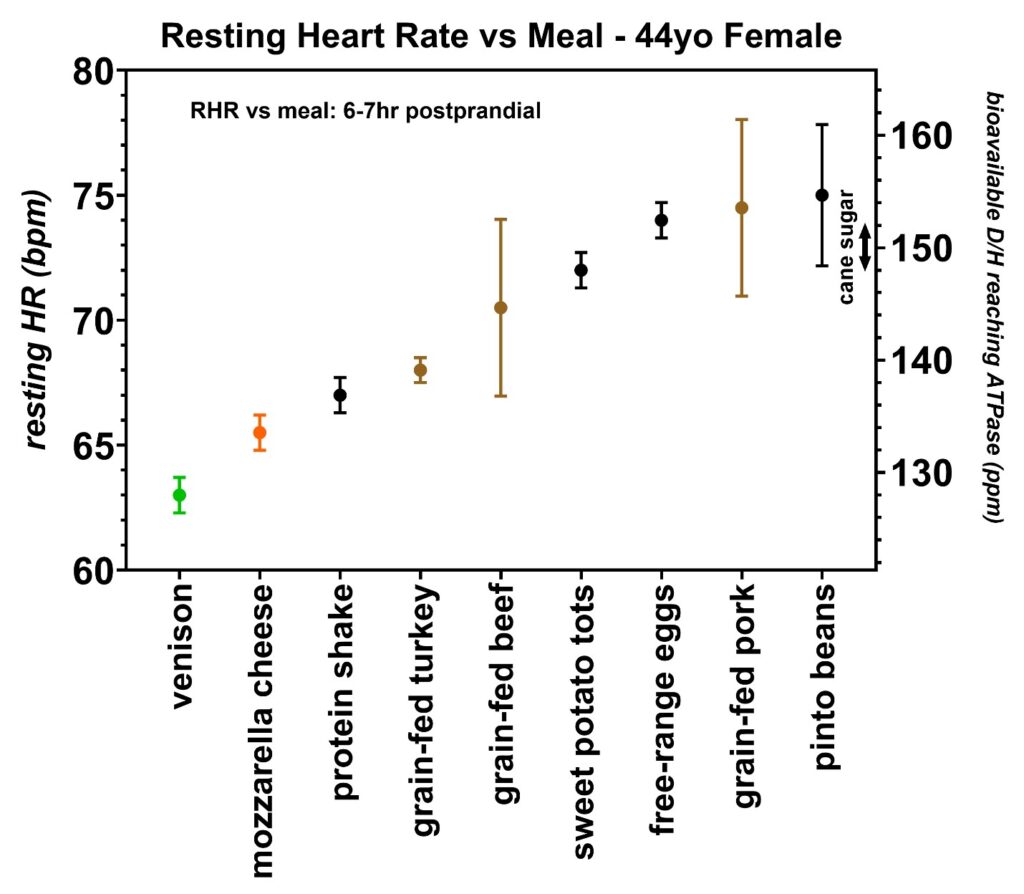 Figure 19: Resting heart rates recorded just prior to awakening over a two-month period collected by volunteer 5 (44-year-old female). This female volunteer tested some naturally-fed venison, grain-fed turkey, grain-fed beef, and grain-fed pork to determine if the resting heart rates varied as greatly as volunteer 2, who was also female. The calibration equation slopes were different in these two volunteers (Figure 6), ruling out sex as the cause of the increased resting heart rate range as observed by volunteer 2. The D/H ratio for cane sugar measured with mass spectroscopy is also indicated.
Figure 19: Resting heart rates recorded just prior to awakening over a two-month period collected by volunteer 5 (44-year-old female). This female volunteer tested some naturally-fed venison, grain-fed turkey, grain-fed beef, and grain-fed pork to determine if the resting heart rates varied as greatly as volunteer 2, who was also female. The calibration equation slopes were different in these two volunteers (Figure 6), ruling out sex as the cause of the increased resting heart rate range as observed by volunteer 2. The D/H ratio for cane sugar measured with mass spectroscopy is also indicated.
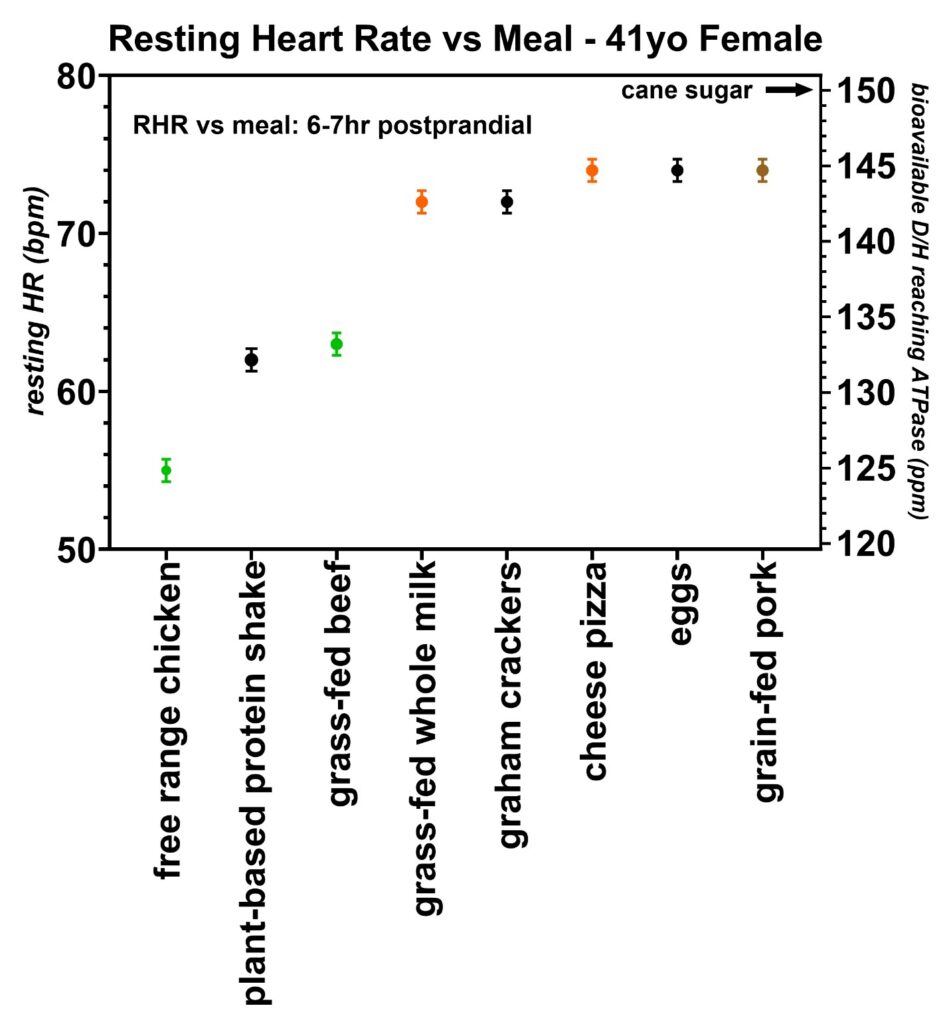 Figure 20: Resting heart rates recorded just prior to awakening over a two-month period from volunteer 6 (41-year-old female). This volunteer tested free-range chicken, grass-fed beef, and grain-fed pork and found the same general trend in increasing resting heart rates with increasing deuterium levels. All six volunteers were found to have unique resting heart rate vs. deuterium level calibrations (Figure 6), as discussed in more detail earlier in the article. The D/H ratio for cane sugar measured with mass spectroscopy is also indicated.
Figure 20: Resting heart rates recorded just prior to awakening over a two-month period from volunteer 6 (41-year-old female). This volunteer tested free-range chicken, grass-fed beef, and grain-fed pork and found the same general trend in increasing resting heart rates with increasing deuterium levels. All six volunteers were found to have unique resting heart rate vs. deuterium level calibrations (Figure 6), as discussed in more detail earlier in the article. The D/H ratio for cane sugar measured with mass spectroscopy is also indicated.
Deuterium content of proteins
Proteins were further tested during the exercise state to determine if they directly feed the TCA cycle in the cardiomyocytes. On two separate hikes up a 1254 m net elevation gain, 40 g of whey protein was consumed 2 h into each hike, as shown in Figure 21. In both instances, the exercise heart rate began decreasing within 15 min of consuming the protein shakes. This is strongly suggestive that the amino acids are directly feeding the TCA cycle of the cardiomyocytes because 15 min is an insufficient amount of time to convert the amino acids into fatty acids for transport into the cardiomyocytes via the CD36 receptor [36]. The decreases in exercise heart rate also imply that these whey protein shakes have a low bioavailable deuterium level. Proteins are known to be transported into cells via several amino acid transporters [42]. Finally, the exact entry point of the TCA cycle depends on the specific amino acid involved, and details of these processes are already well described in metabolism textbooks [42].
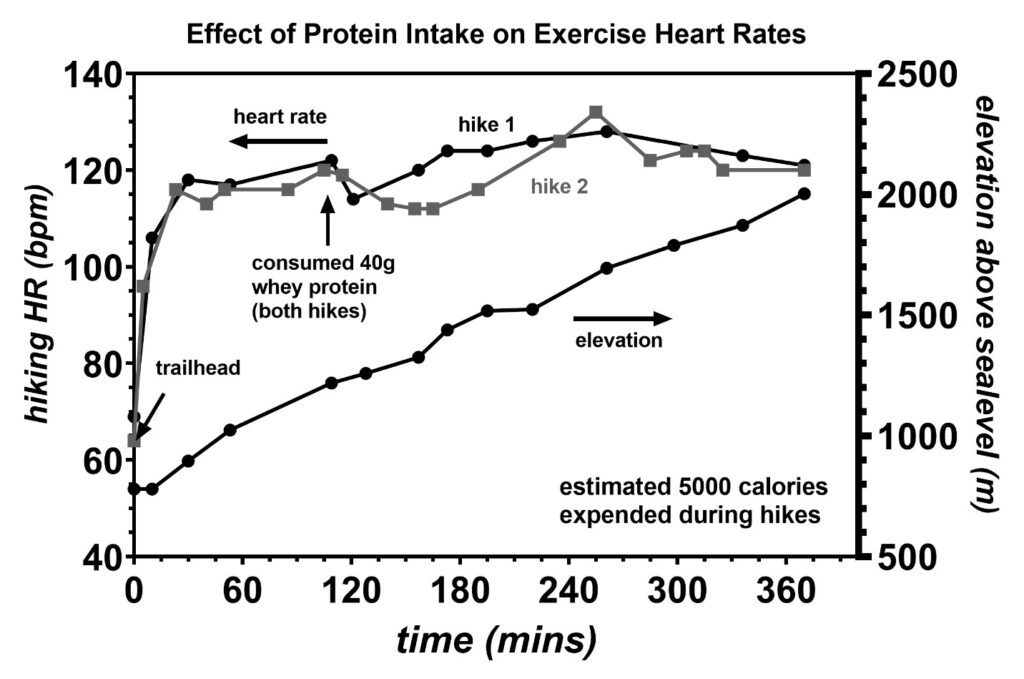 Figure 21: Exercise heart rates recorded during two strenuous hikes up a 2003 m tall mountain or 1254 m net elevation gain. During each hike, a whey protein shake containing 40 g of protein was consumed 2 h into each hike. In both cases, the exercise heart rate was found to decrease within 15 min of consuming these protein shakes indicating that the amino acids were directly feeding the TCA cycle of the cardiomyocytes. Similar to the observation of the effect of protein shakes inducing low resting heart rates, the decrease in the exercise heart rates following protein shake consumption is consistent with the protein shakes having a low bioavailable deuterium level.
Figure 21: Exercise heart rates recorded during two strenuous hikes up a 2003 m tall mountain or 1254 m net elevation gain. During each hike, a whey protein shake containing 40 g of protein was consumed 2 h into each hike. In both cases, the exercise heart rate was found to decrease within 15 min of consuming these protein shakes indicating that the amino acids were directly feeding the TCA cycle of the cardiomyocytes. Similar to the observation of the effect of protein shakes inducing low resting heart rates, the decrease in the exercise heart rates following protein shake consumption is consistent with the protein shakes having a low bioavailable deuterium level.
Mercury exposure from seafoods
Seafoods are among the lowest deuterium-containing foods on the planet. However, some seafoods, when eaten in excess, are known to raise mercury levels in the body [43]. These generally include the fish farther down the food chain. Shark, pike, albacore, and halibut are typically considered the highest mercury-containing fish and are recommended not to be eaten more than once per month. In order to obtain accurate deuterium levels, a minimum of five meals from each of the 30 fish species tested meant at least 150 dinners of testing. As a result, volunteer 1 had hair analysis run twice during this phase of testing to determine if excessive mercury exposures were taking place. These tests shown in Figure 22 were conducted by evenbetternow.com [44]. During the first half of the experiments, when oysters, clams, and warmer water fish were being consumed nearly every evening, the hair mercury level did briefly exceed the recommended maximum value of 0.15 mg%. A level of 0.21 mg% was recorded at the age of 60.70 years. Fortunately, mercury levels decreased below the 0.15 mg% threshold during the latter half of the seafood testing when mostly colder water fish were being consumed nearly every night. It was concluded that consumption of seafoods no more than every other night is unlikely to raise hair levels beyond the 0.15 mg% threshold currently considered safe. Ironically, serum blood tests did not detect any mercury exposure, likely because the heavy metals are rapidly deposited into body tissues soon after exposure.
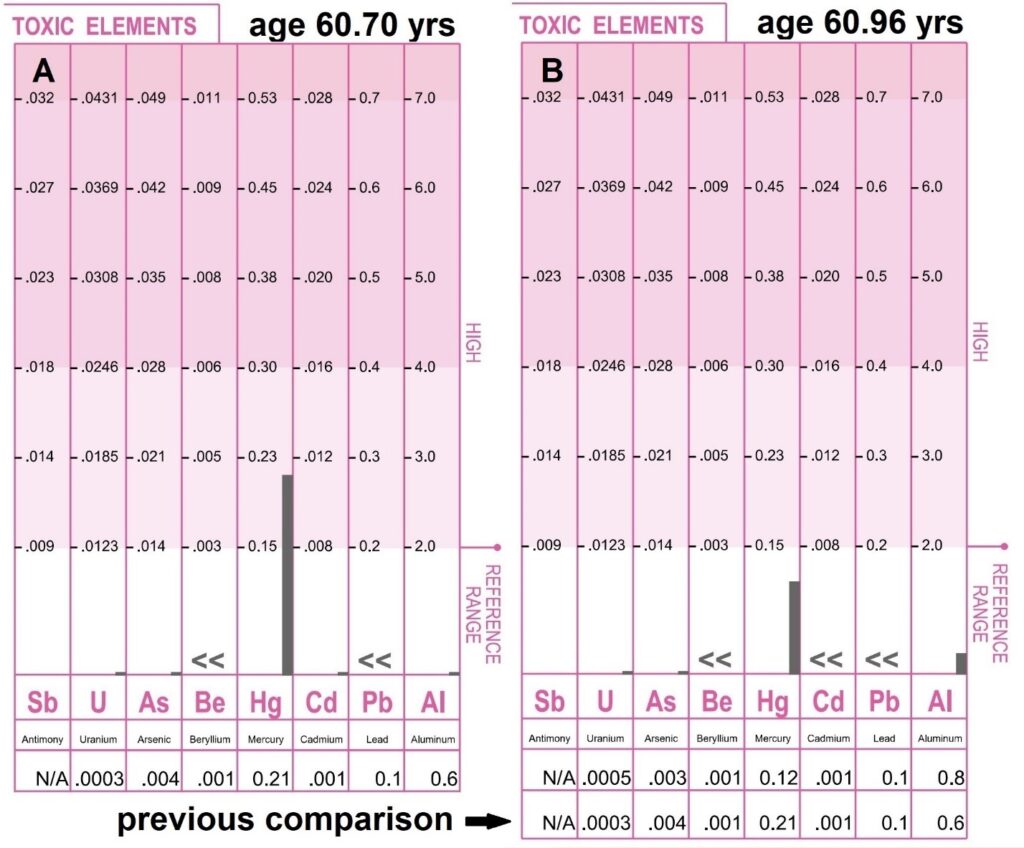 Figure 22: Toxic elements were measured in the scalp hair of volunteer 1 at the age of 60.70 years and 60.96 years during the seafood tests. To obtain more accurate deuterium levels, each seafood was consumed a minimum of five times to improve the accuracy of the resting heart rate average, putting the volunteer at a higher risk of increased mercury exposure [43]. As a result, hair analysis was conducted twice by evenbetternow.com during these seafood tests [44]. Panel A was obtained following a period of high oyster, clam, and warmer water fish consumption. Panel B was a repeat test when the majority of the seafoods consumed were the colder water fish. Toxic elements are reported in units of milligrams percent (milligrams per one hundred grams of hair). One milligram percent (mg%) is equal to ten parts per million (ppm). Values less than 0.15 mg% are considered safe. The mercury level did reach 0.21 mg% for a brief period during the time the warmer water fish were being consumed daily. Ironically, serum blood mercury levels were zero.
Figure 22: Toxic elements were measured in the scalp hair of volunteer 1 at the age of 60.70 years and 60.96 years during the seafood tests. To obtain more accurate deuterium levels, each seafood was consumed a minimum of five times to improve the accuracy of the resting heart rate average, putting the volunteer at a higher risk of increased mercury exposure [43]. As a result, hair analysis was conducted twice by evenbetternow.com during these seafood tests [44]. Panel A was obtained following a period of high oyster, clam, and warmer water fish consumption. Panel B was a repeat test when the majority of the seafoods consumed were the colder water fish. Toxic elements are reported in units of milligrams percent (milligrams per one hundred grams of hair). One milligram percent (mg%) is equal to ten parts per million (ppm). Values less than 0.15 mg% are considered safe. The mercury level did reach 0.21 mg% for a brief period during the time the warmer water fish were being consumed daily. Ironically, serum blood mercury levels were zero.
Discussion
Deuterium has been linked to many medical disorders, including lung [19], rare childhood [20], renal cell [21], colorectal cancers [22], depression [23], diabetes mellitus [24, 25], and heart failure [3]. This is due to deuterium slowing the rate of ATP production, shifting the cellular energy production towards the acidic, anaerobic glycolysis pathway. Furthermore, deuterium is also believed to hinder the repair of DNA in the nuclei [45]. In the heart, it was recently discovered that the heart rate varies systematically with the amount of deuterium contained in the fatty acids and proteins of the foods recently consumed [3]. This is due to deuterium decreasing the cardiac stroke volume, thereby increasing the heart rate needed to maintain an adequate cardiac output. When the heart becomes unable to meet the cardiac output demands, heart failure occurs. If the structural integrity of the heart is intact, HFpEF occurs [3].
Mass spectroscopy has determined that the deuterium content in food is the highest in carbohydrates, followed by fats and proteins [3, 34]. Furthermore, it was also shown both experimentally in 6 volunteers and theoretically that the D/H levels are higher in grain-fed land animals compared to grass-fed land animals. In this report, seafoods caught in the top 500 m of the water are also shown to have a progressively lower D/H value as the food is caught closer to the colder water adjacent to the melting ice. This corresponds to the lower deuterium levels in the colder regions as directly measured in past isotope studies of the oceans published elsewhere [16]. These findings further confirm that the resting heart rates are clearly tracking the deuterium levels in the fatty acid and protein composition of the foods eaten. It is interesting to point out here that the omega-3/omega-6 ratios are also recognized to be higher in the seafoods caught from colder waters [46], and although unproven here, it is plausible that deuterium could be slowing the production of the more frequent double bonds needed in the biological production of the anti-inflammatory omega-3 fatty acids.
Artificial supplements such as vitamins are manufactured with Vienna Standard hydrogen, i.e., 155.76 ppm D/H, which is likely higher than their natural counterparts obtained directly from plants. Furthermore, partial hydrogenation of oils to increase the shelf life and/or increase their viscosity is also most likely done with the Vienna Standard hydrogen making these fats less healthy. The more frequent deuterium bonds in these partially hydrogenated oils are possibly connected to the mechanism causing the trans-fat configuration to occur.
Our bodies have two mechanisms for eliminating deuterium from the metabolic substrates prior to them entering the mitochondria in order to protect the ATP nanomotors from the detrimental effect of this deuterium. First, glucose from dietary carbohydrate intake enters a 10-step glycolysis process to produce two pyruvates that get transported into the mitochondria [47]:
Glucose + 2 NAD+ + 2 ADP + 2 Pi –> 2 Pyruvate + 2 NADH + 2 H+ + 2 ATP + 2 H2O.
As shown in Figure 23, a net loss of four hydrogens which are relatively high in deuterium ratio (D/H ~ 150 ppm in glucose), are removed during this process prior to the pyruvates being transported across the mitochondria membrane. This, in essence, is a “deuterium drying” process. Unfortunately, when carbohydrates are consumed in excess, a spike in insulin occurs, leading to glucose entering the adipose tissue where it is converted into a high deuterium storage fat. When these high D/H fats are ultimately used to produce the cellular energy in the mitochondria via the beta-oxidation process, this “deuterium drying” process is bypassed. This, in turn, severely limits the efficiency of the ATP nanomotors [47]. Second, the data presented below suggests that the enzyme adipose triglyceride lipase (ATGL) releases higher deuterium triglycerides into blood circulation at a faster rate than the lower deuterium triglycerides. Although a limited amount of information is known about this ATGL enzyme, this enzyme is known to be regulated by a minimum of six proteins, two small molecules, and pH [48]. The peak activity of ATGL occurs at pH 7.0 with a decrease to 50% activity level at 1 pH unit to either side of pH 7.0. One small regulatory molecule includes long-chain acyl-CoAs which inhibit the ATGL activity [48]. It is possible that the ATGL activity could also be influenced by a deuterium hydrogen isotope effect. In addition, it is possible that deuterium could be more concentrated in the shorter chain acyl-CoAs, which would ultimately lead to the ATGL enzyme releasing the higher D/H fatty acids into blood circulation faster than the lower D/H fatty acids. The fasting resting heart rate data shows that the adipose tissue is being converted into a deuterium-depleted energy pool with physical activity. This process is most pronounced with frequent exercise, where the skeletal muscle eliminates the unhealthy high deuterium triglycerides faster, allowing the healthier low D/H triglycerides to remain behind for a more efficient energy supply for the organs. Furthermore, this effect is also readily noticed when an individual first begins to exercise. Most individuals typically report a sluggish performance at the onset of exercise which progressively improves as time advances. The initial sluggish response is due to the higher deuterium triglycerides being released to the muscles first. It is also one of the reasons why warm-ups are important prior to sporting events.
When the skeletal fat deuterium levels become excessive, either by an inadequate amount of exercise and/or by high intake of high deuterium-containing foods, cardiac stroke volume decreases. This then leads to two cardiovascular compensating mechanisms that are needed to maintain adequate cardiac outputs. First, the decrease in stroke volume causes a secondary increase in autonomic vascular tone to help maintain blood pressure [3]. Any decrease in blood pressure from an incomplete vascular response will then lead to a secondary compensating response by the heart. The SA node acting as a baroreceptor will increase the heart rate to maintain the needed cardiac output [3, 36]. When both compensation mechanisms are insufficient to meet the metabolic demands of the body and blood pressure requirements, heart failure is said to occur. Therefore, for individuals with good compensating mechanisms, the body is able to tolerate higher levels of dietary deuterium intakes without heart failure occurring. In the individuals with good cardiac functioning but decreased vascular tone, the rate of increase in the heart rate is larger than those with the good vascular tone, and an example of this scenario was observed in volunteer 2 (Figure 15), who has a long history of SLE [35].
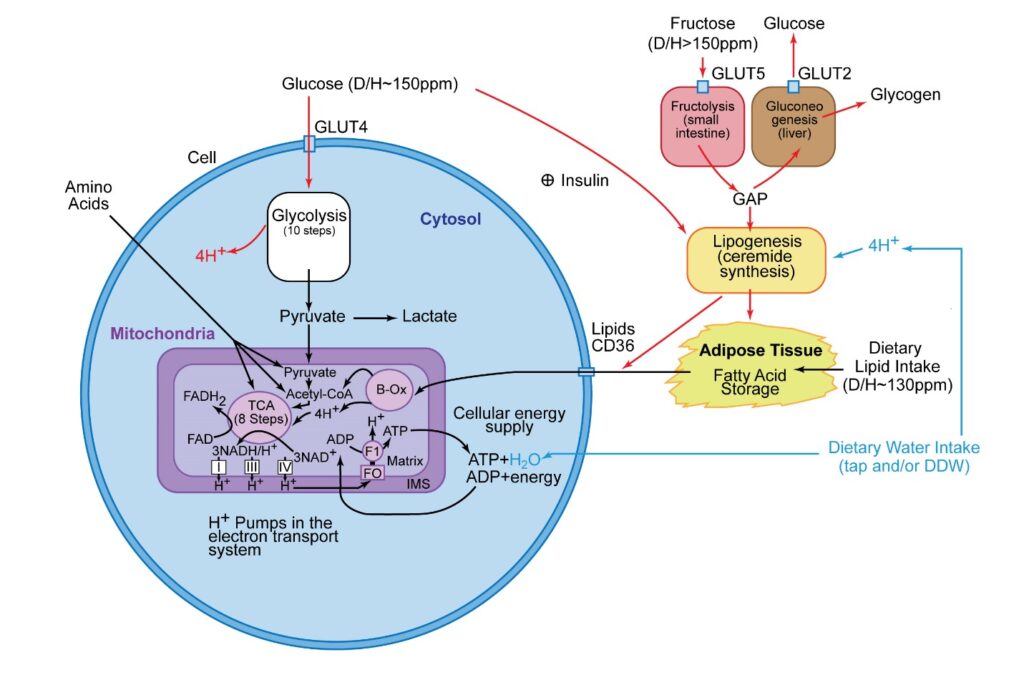 Figure 23: The basic metabolic energy pathways involved in the production of ATP in a typical cell. In this figure, the higher deuterium pathways are shown as red arrows, the lower deuterium pathways are shown as black arrows, and the water intake pathways are shown as blue arrows. In the cardiomyocytes, the primary energy pathway is via lipids, secondarily followed by amino acids. The glycolysis steps in cardiomyocytes only occur during anaerobic metabolism induced during myocardial infarctions. In most cells outside the heart, pyruvate production occurs in the cytoplasm via a 10-step glycolysis process that strips off 4 hydrogens that are relatively high in deuterium, creating a “deuterium drying” process for this energy substrate [47]. This ultimately enters the mitochondria, where the lower deuterium substrate protects the ATP nanomotors from the detrimental effect of high deuterium. Unfortunately, when carbohydrates are consumed in excess, an insulin spike loads the higher deuterium glucose into the adipose tissue, where it is converted into high deuterium fat. This high D/H fat can later enter a cell via the CD36 receptor, which bypasses this “deuterium drying” process. This ultimately slows the ATP nanomotor rotation, decreasing the peak rate of ATP production. This ATP nanomotor slowdown has been linked to a decrease in cardiac stroke volume, which increases the heart rate [3] seen in the data presented in this report. When the deuterium levels increase to the threshold where the cardiac output can no longer be sustained by a compensating increase in heart rate, heart failure occurs [3]. It is important to point out that fructose is the highest deuterium-containing substrate found in food (D/H > 150 ppm), and fructose is rapidly converted to very high deuterium storage fat without the need for insulin. Typical fats from healthy food sources average ~130–140 ppm D/H as compared to fats produced from glucose and high fructose corn syrups (HFCS) (D/H ~150–166 ppm) as measured from mass spectroscopy. Amino acids can also serve as substrates to pyruvate, acetyl-CoA, or the TCA cycle, and the entry site is dependent on the specific amino acid involved [42]. Under aerobic conditions, pyruvate enters the TCA cycle and undergoes oxidative phosphorylation leading to a net production of 32 ATP molecules. Under anaerobic conditions or high mitochondrial deuterium levels, which stall the ATP nanomotors, pyruvate converts to lactate through an anaerobic glycolysis pathway in the cytoplasm, only resulting in the net production of 2 ATP molecules. This process occurs during myocardial infarction, and lactate is likely the trigger involved in the etiology of classic chest pain. Figure adopted from reference 3.
Figure 23: The basic metabolic energy pathways involved in the production of ATP in a typical cell. In this figure, the higher deuterium pathways are shown as red arrows, the lower deuterium pathways are shown as black arrows, and the water intake pathways are shown as blue arrows. In the cardiomyocytes, the primary energy pathway is via lipids, secondarily followed by amino acids. The glycolysis steps in cardiomyocytes only occur during anaerobic metabolism induced during myocardial infarctions. In most cells outside the heart, pyruvate production occurs in the cytoplasm via a 10-step glycolysis process that strips off 4 hydrogens that are relatively high in deuterium, creating a “deuterium drying” process for this energy substrate [47]. This ultimately enters the mitochondria, where the lower deuterium substrate protects the ATP nanomotors from the detrimental effect of high deuterium. Unfortunately, when carbohydrates are consumed in excess, an insulin spike loads the higher deuterium glucose into the adipose tissue, where it is converted into high deuterium fat. This high D/H fat can later enter a cell via the CD36 receptor, which bypasses this “deuterium drying” process. This ultimately slows the ATP nanomotor rotation, decreasing the peak rate of ATP production. This ATP nanomotor slowdown has been linked to a decrease in cardiac stroke volume, which increases the heart rate [3] seen in the data presented in this report. When the deuterium levels increase to the threshold where the cardiac output can no longer be sustained by a compensating increase in heart rate, heart failure occurs [3]. It is important to point out that fructose is the highest deuterium-containing substrate found in food (D/H > 150 ppm), and fructose is rapidly converted to very high deuterium storage fat without the need for insulin. Typical fats from healthy food sources average ~130–140 ppm D/H as compared to fats produced from glucose and high fructose corn syrups (HFCS) (D/H ~150–166 ppm) as measured from mass spectroscopy. Amino acids can also serve as substrates to pyruvate, acetyl-CoA, or the TCA cycle, and the entry site is dependent on the specific amino acid involved [42]. Under aerobic conditions, pyruvate enters the TCA cycle and undergoes oxidative phosphorylation leading to a net production of 32 ATP molecules. Under anaerobic conditions or high mitochondrial deuterium levels, which stall the ATP nanomotors, pyruvate converts to lactate through an anaerobic glycolysis pathway in the cytoplasm, only resulting in the net production of 2 ATP molecules. This process occurs during myocardial infarction, and lactate is likely the trigger involved in the etiology of classic chest pain. Figure adopted from reference 3.
During the fasting state, the fats in circulation are no longer dominated by those consumed in the most recent meal but rather those released from the skeletal fat. Therefore, the fasting resting heart rate serves as a sensitive probe of the level of deuterium in our stored skeletal fat [3]. Volunteer 1 recorded fasting resting heart rates on the occasions when he was able to get between 11 and 14 h of sleep during the 20-month testing period. Figure 24 shows his fasting resting heart rates between the age of 59.6 and 61.3 years with the estimated D/H obtained from the calibration equation obtained from Figure 6. The general activity levels during several time segments are shown. The effect of three strenuous backpacking trips on the fasting resting heart rates is also indicated. On other occasions, the delayed effects of blueberries, eggnog (containing some high fructose corn syrup), and grain-fed pork are also indicated. The findings reveal that significant drops in the skeletal deuterium levels occur following strenuous backpacking trips indicating the higher deuterium fats are more rapidly utilized by the muscles during this strenuous activity. In contrast, increases in the skeletal fat deuterium levels are observed following the consumption of fructose. This is due to the rapid conversion of these higher deuterium carbohydrates into storage fats [37], as shown in the metabolic pathways in Figure 23. The most important finding revealed by these data is that the deuterium level in the skeletal fat can drop lower than those induced by any of the individual foods consumed. This strongly suggests that the ATGL enzyme [48] is releasing the higher deuterium triglycerides into the bloodstream at a faster rate than the lower deuterium triglycerides. It is also the reason that some endurance athletes have recorded resting heart rates as low as 27 bpm [28]! Finally, there were brief increases in the adipose tissue deuterium levels following two asymptomatic COVID-19 infections. These viruses were previously reported to infect the adipose tissue [49], and the mechanism for these changes is unclear. However, deuterium was reported by others to stabilize the viral protein and protect them from temperature denaturation [26].
 Figure 24: Fasting resting heart rates from volunteer 1 between the age of 59.6 and 61.3 years. General activity levels given as km hiked per week are indicated. Furthermore, three strenuous backpacking trips are also indicated showing drops in the fasting resting heart rates following these trips. In contrast, increase in fasting resting heart rates following the consumption of fructose and/or high fructose corn syrup (HFCS) which are rapidly converted into high deuterium skeletal storage fats [37]. Following two of the three backpack trips, the fasting resting heart rate actually dropped lower than any of the individual food-induced resting heart rates indicating that the adipose triglyceride lipase (ATGL) enzyme [48] is mobilizing higher D/H skeletal fats at a faster rate than the lower D/H skeletal fats as discussed in the article. Regular exercise thus converts skeletal fat into a low deuterium triglyceride energy pool. There were brief increases in skeletal fat deuterium levels following two asymptomatic COVID-19 Omicron infections indicating that this virus does infect adipose tissue cells as reported by other researchers [49]. There was also a brief increase in the fasting resting heart rate following some consumption of grain-fed pork as indicated.
Figure 24: Fasting resting heart rates from volunteer 1 between the age of 59.6 and 61.3 years. General activity levels given as km hiked per week are indicated. Furthermore, three strenuous backpacking trips are also indicated showing drops in the fasting resting heart rates following these trips. In contrast, increase in fasting resting heart rates following the consumption of fructose and/or high fructose corn syrup (HFCS) which are rapidly converted into high deuterium skeletal storage fats [37]. Following two of the three backpack trips, the fasting resting heart rate actually dropped lower than any of the individual food-induced resting heart rates indicating that the adipose triglyceride lipase (ATGL) enzyme [48] is mobilizing higher D/H skeletal fats at a faster rate than the lower D/H skeletal fats as discussed in the article. Regular exercise thus converts skeletal fat into a low deuterium triglyceride energy pool. There were brief increases in skeletal fat deuterium levels following two asymptomatic COVID-19 Omicron infections indicating that this virus does infect adipose tissue cells as reported by other researchers [49]. There was also a brief increase in the fasting resting heart rate following some consumption of grain-fed pork as indicated.
It is useful to summarize the energy substrate pathways at this point, separating the pathways that raise the deuterium levels from those that lower the deuterium levels. From mass spectroscopy, plants are known to move higher levels of deuterium toward the plant storage starches [34]. As a result, carbohydrates tend to supply energy substrates with deuterium levels of D/H ~ 150 ppm. In contrast, most fats and oils have lower D/H ~ 130–140 ppm. Furthermore, proteins generally fall into the range of fats with D/H ~ 130–140 ppm [3, 34]. Figure 25 illustrates these general energy pathways in our bodies. The crucial energy pool for our heart is the storage fat labeled “body fat”. When this fat becomes contaminated with high deuterium triglycerides from either overconsumption of carbohydrates or insufficient levels of exercise, the heart stroke volume decreases. This can lead to an exacerbation of heart failure or the development of heart failure in medically compromised individuals. The enzyme ATGL [48] mobilizes the higher deuterium fats at a faster rate than the lower deuterium fats converting the skeletal fat stores into a low deuterium energy pool with regular frequent exercise. This enzyme is also the reason that athletes initially feel sluggish at the onset of their exercise session, with their energy level gradually increasing throughout the session because the higher D/H fats are utilized by the muscle first. These higher D/H fats are unable to produce ATP at the maximal rate causing this initial sluggish feeling. However, once the deuterons are passed through the ATP nanomotors, they bind to oxygen-forming water which eventually gets transported into the blood plasma. There the higher deuterium water can be excreted via urine or sweat. The deuterium can also be diluted by consumption of DDW, assisting in the removal of this deuterium from the body.
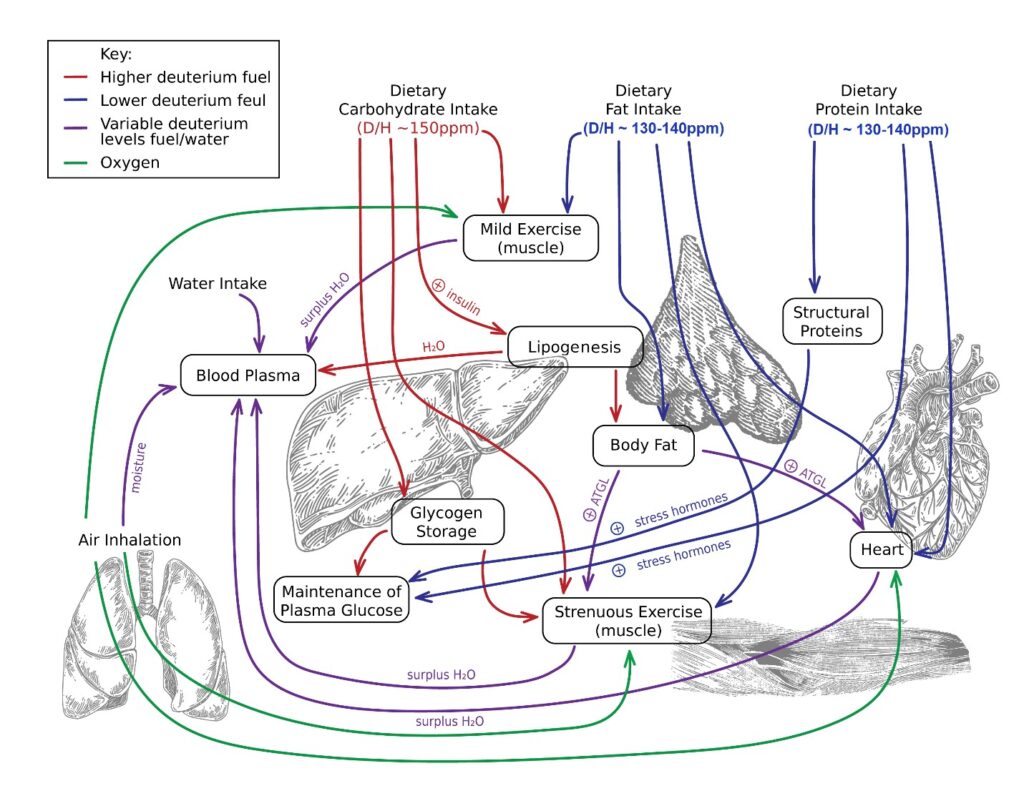 Figure 25: Movement pathways for the energy substrates encoded according to their relative deuterium concentrations. The carbohydrates are generally high in deuterium (~150 ppm D/H) based on mass spectroscopy studies [3, 34]. These pathways are encoded as red arrows. The fats and proteins are generally low in deuterium (~ 130–140 ppm D/H) [3, 34], and these are encoded as blue arrows. Oxygen pathways to the organs are encoded as green arrows, and these pathways are included in the chart here because this oxygen binds to the hydrogen after they pass through the ATP nanomotors leading to the production of water. The water produced by this process can be either high or low in deuterium and this water ultimately transfers into the blood plasma. Due to the variable D/H levels in the water, water is encoded as purple arrows. The most significant part of this plot is the production of storage fat labeled as “body fat”. Lipogenesis from excessive carbohydrate intake produces high deuterium storage fat (labeled red), whereas direct storage of fat from dietary intake of fat is lower in deuterium (labeled as blue). The adipose triglyceride lipase (ATGL) enzyme [48, 50, 51] mobilizes the higher deuterium fat at a faster rate than the lower deuterium fat. As discussed in the text, the ATGL activity is regulated by six known proteins, two small molecules, and pH [48]. It may also be influenced by a deuterium hydrogen isotope effect. Regular strenuous exercise over time will lower the deuterium level of the storage fat leading to an increase in cardiac stroke volume and lowering the heart rate needed to maintain the cardiac output. When the deuterium levels become excessive in the storage fat, the cardiac stroke volume decreases, thereby increasing the heart rate [3]. This is exacerbated if some of the cardiac muscle has already been damaged and replaced with fibrotic tissue [36]. When the cardiac conduction system is unable to maintain an adequate heart rate, heart failure will occur.
Figure 25: Movement pathways for the energy substrates encoded according to their relative deuterium concentrations. The carbohydrates are generally high in deuterium (~150 ppm D/H) based on mass spectroscopy studies [3, 34]. These pathways are encoded as red arrows. The fats and proteins are generally low in deuterium (~ 130–140 ppm D/H) [3, 34], and these are encoded as blue arrows. Oxygen pathways to the organs are encoded as green arrows, and these pathways are included in the chart here because this oxygen binds to the hydrogen after they pass through the ATP nanomotors leading to the production of water. The water produced by this process can be either high or low in deuterium and this water ultimately transfers into the blood plasma. Due to the variable D/H levels in the water, water is encoded as purple arrows. The most significant part of this plot is the production of storage fat labeled as “body fat”. Lipogenesis from excessive carbohydrate intake produces high deuterium storage fat (labeled red), whereas direct storage of fat from dietary intake of fat is lower in deuterium (labeled as blue). The adipose triglyceride lipase (ATGL) enzyme [48, 50, 51] mobilizes the higher deuterium fat at a faster rate than the lower deuterium fat. As discussed in the text, the ATGL activity is regulated by six known proteins, two small molecules, and pH [48]. It may also be influenced by a deuterium hydrogen isotope effect. Regular strenuous exercise over time will lower the deuterium level of the storage fat leading to an increase in cardiac stroke volume and lowering the heart rate needed to maintain the cardiac output. When the deuterium levels become excessive in the storage fat, the cardiac stroke volume decreases, thereby increasing the heart rate [3]. This is exacerbated if some of the cardiac muscle has already been damaged and replaced with fibrotic tissue [36]. When the cardiac conduction system is unable to maintain an adequate heart rate, heart failure will occur.
The ATGL enzyme was recognized as vital for maintaining a balance between lipid storage and mobilization in 2009 [50]. Dysregulated lipolysis was recognized as playing a role in metabolic diseases, including insulin resistance, diabetes, inflammation, and non-alcoholic fatty liver [48, 50]. By 2020, the lipolytic processes in adipose tissue modulated by the ATGL activity were also reported to play a role in the development of heart failure, in particular, HFpEF which accounts for 2/3 of the heart failure cases [51]. It was theorized that pharmacological inhibition of the ATGL enzyme might serve as a future treatment option for heart failure [51]. However, the underlying pathology is actually a slow accumulation of the higher deuterium-containing lipids inside the adipose tissue which ultimately reach and stall the ATP nanomotors slowing ATP production [9].
Excessive visceral fat is generally considered to be more harmful to health than excessive skeletal fat. This is because visceral fat coats the internal organs of the portal circulation. Furthermore, there is some evidence that visceral fat might be more inflammatory than skeletal fat [52]. It is useful to comment here that the pH of the portal blood circulation has never been characterized. Not a single portal blood pH value has ever been located in the medical literature. Depending on the diet of an individual, the portal blood pH may not necessarily match the pH of the systemic blood which is 7.4. Furthermore, the cytoplasm pH of the visceral fat cells may be well off the peak activity level of ATGL, i.e., pH 7.0 [48], that some individuals are unable to utilize the visceral fat stores as rapidly as the skeletal fat stores causing these individuals to slowly accumulate visceral fat with age. However, since the portal blood pH has not been characterized, there is no definitive answer as to whether or not this may be occurring in these individuals.
There are other aspects of the cardiovascular system that are influenced by deuterium levels. There are at least five points from the CHA2DS2-VASc stroke risk score in atrial fibrillation [53, 54] that are attributable to high deuterium levels. Congestive heart failure contributes one point to the total score, and this topic is the focus of this report. Age of 75 years or older contributes two points to the score, and an increase in deuterium is expected with the slowing of physical activity with age. The slowing of physical activity allows the deuterium levels to accumulate in the skeletal fat, and this issue is also discussed in this report. Diabetes mellitus also contributes one point to the total score, and the connection of diabetes to deuterium levels is published elsewhere [24, 25]. Vascular disease is an inflammatory condition with elevated CRP levels [38], and CRP was found to decrease with the consumption of DDW in volunteer 1 (Figure 7). The vascular disease also contributes one additional point to the total score. The contribution to the total score from other conditions, hypertension (one point), previous stroke/TIA (two points), and female gender (one point), have unclear connections to the deuterium levels at this time. However, the five points out of a maximum score of nine points listed here already suggest that high plasma deuterium levels may significantly increase stroke risk.
Elevated heart rates can also be found with the use of ethanol [55]. In this substance, the deuterium is stable in the alcohol molecule and this molecule is produced by the fermentation of high deuterium sugars, rice, corn, and potato starches. Interestingly, volunteer 4 did record a relatively higher resting heart rate following a light consumption of wine (Figure 18). The alcohol molecule is known to target the GABA pathways in the brain where the deuterium likely slows the production rate of ATP in those regions leading to the relaxation and/or intoxication effects of the alcohol. During the hepatic clearance of the alcohol from the body, some of the detrimental toxicity is likely caused by the slowing of ATP production in the hepatocytes needed to clear the alcohol from the system. It is interesting to point out that researchers in China have studied the effect of deuterium-depleted alcohol in lab rats to better elucidate these cellular damage mechanisms [56].
Some individuals are especially sensitive to the toxic effects of high deuterium levels. These individuals include the medically ill who cannot perform regular exercise to activate their ATGL enzyme [48, 50, 51] to mobilize the high D/H triglycerides for use in their skeletal muscles. Space explorers are also susceptible due to them being weightless, which severely limits their ability to mobilize these high D/H triglycerides via the ATGL enzyme [48, 50, 51] for their skeletal muscles. These individuals need to be especially careful with their consumption of higher D/H foods. The Astronaut Twin Study does show that astronauts enter an anaerobic, lactic acid-producing energy pathway shortly after becoming weightless [57]. Their urine shows increases in lactic acid levels while in orbit [57]. In medically ill hospitalized patients, the introduction of deuterium-depleted IV fluids could prove extremely beneficial to their prognosis. However, due to the weight limitations, space explorers are more likely to benefit from the consumption of low-deuterium foods, which could include daily consumption of olive and/or avocado oil for the production of low-deuterium metabolic water in their mitochondria. It is important to note that high levels of deuterium are believed to increase the susceptibility to radiation expected in space.
Conclusion
Heart failure results when the cardiac output becomes insufficient to meet the demands of the body [1, 2]. It can occur as a result of structural damage and/or a decrease in the ability to rapidly produce ATP cellular energy. In the cases of heart failure where the structural integrity remains intact, the ejection fraction (EF) remains normal. This subtype of heart failure is called HFpEF. Data presented in this report show that the ability of the cells to produce ATP cellular energy is slowed with a progressive increase in the consumption of high deuterium-containing foods. These foods often include high carbohydrate content, fructose, and seed oils. Consumption of these foods is found to lead to higher heart rates due to a corresponding decrease in cardiac stroke volume. Therefore, when the heart rates are unable to be maintained by the finite speed of the heart conduction system, heart failure appears. Furthermore, the consumption of these same foods is also believed to contribute to higher cancer rates seen in many countries [47].
Healthy individuals who are able to exercise on a regular basis are able to eliminate these higher deuterium fats due to the ATGL enzyme [48, 50, 51] mobilizing these higher D/H triglycerides to the skeletal muscle at a faster rate than the lower D/H triglycerides. In these healthy individuals, during exercise, the ATGL enzyme converts the skeletal fats into a low deuterium triglyceride energy pool. This low deuterium energy pool allows athletes to develop very low resting heart rates [28]. Furthermore, this residual energy pool also provides for a more efficient healthier energy source for the organs. However, the medically compromised population or space explorers in a weightless environment [57] are unable to eliminate these high D/H triglycerides from the skeletal fat allowing the deuterium levels in the triglyceride stores to slowly increase. This places these individuals at higher risk of not only heart failure but also cancer [19–22, 47], metabolic disorders [24, 25, 40], and immune dysfunction [26, 58]. This population will benefit the most from a strict consumption of low-deuterium food. Furthermore, hospitalized patients might benefit from introducing deuterium-depleted IV fluids.
Studies to date have been made in six individuals with no cardiac dysfunction. This was done to better develop an understanding of the specific impact of deuterium on the human heart. This study reveals that the deuterium load lowers the maximal rate of ATP production, decreasing the cardiac stroke volume [3]. The knowledge gained from these studies can be applied to cardiac-compromised individuals and space explorers. It is likely that deuterium may also be strongly correlated to the CHA2DS2-VASc stroke risk score [53, 54]. Proposing such studies should not pose any significant ethical issues with the institute review boards. Pharmaceutical companies could also produce new products, including deuterium-depleted IV fluids, to better protect the ATP energy production capacity of medically compromised hospitalized individuals. In any event, the deuterium patterns in the foods presented here in this report should be useful for protecting both the healthy and medically compromised population.
The limitations of this study stem from the fact that all foods produce multiple metabolites. Furthermore, mass spectroscopy analysis is a very slow process making it difficult to measure a large number of foods in a single study. Simple foods are known to contain thousands of metabolites, many of which could act on the autonomic nervous system modulating the heart rate [59]. Mass spectroscopy does not identify which chemical constituent in a specific food actually contains the deuterium. Fortunately, the heart rate comparisons between the grass-fed and grain-fed meats do suggest that the differences in the isotope concentrations are the likely mechanism of the different heart rates reported in this study [13].
Patient Consent
A written informed consent was obtained from the six volunteers for the publication of this report.
Funding
This research did not receive grant funding from agencies in the public, commercial, or not-for-profit sectors.
Conflicts of Interest
The authors declare no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Disclaimer
The views expressed in this article are those of the authors and do not necessarily reflect the position or policy of the Veterans Health Administration, Department of Veterans Affairs, or the US Government.
Acknowledgments
Research began on this subject while the lead author was employed by the Oak Ridge National Laboratory and completed while employed by the Department of Veterans Affairs, and was a privately funded research separate from both of these organizations’ funding and time. The authors thank László Boros, Stephen K. Souther, Christina Asberry, Trinity Patty, Brian J. Rollick, James R. Thompson, and David K. Christen for informative discussions.
References
- National Heart, Lung, and Blood Institute. What Is Heart Failure? 2022.
- Hsu S, Fang JC, Borlaug BA. Hemodynamics for the Heart Failure Clinician: A State-of-the-Art Review. J Card Fail. 2022;28(1):133-48.
- Jones EC, Phelps JE, Jardet CL. Cardiac Heart Rate Dependence on Mitochondrial Deuterium 2H Content. Medical Research Archives. 2022;10(5):1-24.
- Rosano GMC, Vitale C, Spoletini I. Metabolic approach to heart failure: the role of metabolic modulators. Egyptian Heart J. 2015;67(3):177-81.
- Meagher P, Adam M, Civitarese R, et al. Heart Failure With Preserved Ejection Fraction in Diabetes: Mechanisms and Management. Can J Cardiol. 2018;34(5):632-43.
- Zhou B, Tian R. Mitochondrial dysfunction in pathophysiology of heart failure. J Clin Invest. 2018;128(9):3716-26.
- Kumar AA, Kelly DP, Chirinos JA. Mitochondrial Dysfunction in Heart Failure With Preserved Ejection Fraction. Circulation. 2019;139(11):1435-50.
- Bui AL, Horwich TB, Fonarow GC. Epidemiology and risk profile of heart failure. Nat Rev Cardiol. 2011;8(1):30-41.
- Olgun A. Biological effects of deuteronation: ATP synthase as an example. Theor Biol Med Model. 2007;4:9.
- Dorgan LJ, Schuster SM. The effect of nitration and D2O on the kinetics of beef heart mitochondrial adenosine triphosphatase. J Biol Chem. 1981;256(8):3910-16.
- Urbauer JL, Dorgan LJ, Schuster SM. Effects of deuterium on the kinetics of beef heart mitochondrial ATPase. Arch Biochem Biophys. 1984;231(2):498-502.
- Dorgan LJ, Urbauer JL, Schuster SM. Metal dependence and thermodynamic characteristics of the beef heart mitochondrial adenosine triphosphatase. J Biol Chem. 1984;259(5):2816-21.
- JLech JC, Dorfsman SI, Répás Z, et al. What to feed or what not to feed-that is still the question. Metabolomics. 2021;17(12):102.
- Semiheavy water. 2022.
- Reference materials for stable isotope analysis. 2023.
- Ferronsky VI, Polyakov VA. Isotopes of the Earth’s Hydrosphere. Dordrecht: Springer; 2012. Chapter 3: Isotopic Composition of Ocean Water; p. 49-65.
- Chirieri E, Arcescu I, Ganea C, et al. The effect of deuteration on the frog retina bioelectrogenesis. Naturwissenschaften. 1977;64(3):149-50.
- The Nobel Prize. The Nobel Prize in Chemistry 1997. 1997.
- Hao D, Sarfaraz MO, Farshidfar F, et al. Temporal characterization of serum metabolite signatures in lung cancer patients undergoing treatment. Metabolomics. 2016;12:58.
- Gros SJ, Holland-Cunz SG, Supuran CT, et al. Personalized Treatment Response Assessment for Rare Childhood Tumors Using Microcalorimetry-Exemplified by Use of Carbonic Anhydrase IX and Aquaporin 1 Inhibitors. Int J Mol Sci. 2019;20(20):4984.
- Pandey N, Lanke V, Vinod PK. Network-based metabolic characterization of renal cell carcinoma. Sci Rep. 2020;10:5955.
- Arima K, Lau MC, Zhao M, et al. Metabolic Profiling of Formalin-Fixed Paraffin-Embedded Tissues Discriminates Normal Colon from Colorectal Cancer. Mol Cancer Res. 2020;18(6):883-90.
- Strekalova T, Evans M, Chernopiatko A, et al. Deuterium content of water increases depression susceptibility: the potential role of a serotonin-related mechanism. Behav Brain Res. 2015;277:237-44.
- Somlyai G, Somlyai I, Fórizs I, et al. Effect of Systemic Subnormal Deuterium Level on Metabolic Syndrome Related and other Blood Parameters in Humans: A Preliminary Study. Molecules. 2020;25(6):1376.
- Jones EC, Jardet CL. Functional Improvement in β-Islet Cells and Hepatocytes with Decreasing Deuterium from Low Carbohydrate Intake in a Type-II Diabetic. Medical Research Archives. 2021;9(6):1-19.
- Mao J, Liu Z, Tang C, et al. Evidence for hydrogen-deuterium exchange in viral particles. Chin Sci Bull. 2004;49:253-57.
- Rossow LM, Fukuda DH, Fahs CA, et al. Natural bodybuilding competition preparation and recovery: a 12-month case study. Int J Sports Physiol Perform. 2013;8(5):582-92.
- Guinness World Records. Lowest heart rate.
- Okura H, Takada Y, Yamabe A, et al. Age- and gender-specific changes in the left ventricular relaxation: a Doppler echocardiographic study in healthy individuals. Circ Cardiovasc Imaging. 2009;2(1):41-46.
- Jensen MT. Resting heart rate and relation to disease and longevity: past, present and future. Scand J Clin Lab Invest. 2019;79(1-2):108-16.
- Biga LM, Bronson S, Dawson S, et al. Anatomy & Physiology. 1st ed. Houston: OpenStax. Chapter 19.4: Cardiac Physiology.
- Merced-Morales A, Daly P, Abd Elal AI, et al. Influenza Activity and Composition of the 2022–23 Influenza Vaccine – United States, 2021–22 Season. MMWR Morb Mortal Wkly Rep. 2022;71:913-19.
- Valentini M, Parati G. Variables influencing heart rate. Prog Cardiovasc Dis. 2009;52(1):11-19.
- Deuterium Content in different foods and Deuterium Depletion. 2020.
- Barile-Fabris L, Hernández-Cabrera MF, Barragan-Garfias JA. Vasculitis in systemic lupus erythematosus. Curr Rheumatol Rep. 2014;16(9):440.
- Martínez MS, García A, Luzardo E, et al. Energetic metabolism in cardiomyocytes: molecular basis of heart ischemia and arrhythmogenesis. Vessel Plus. 2017;1:130-41.
- Keim NL, Havel PJ. Encyclopedia of Human Nutrition. 3rd ed. Waltham: Academic Press; 2013. Fructose: Absorption and Metabolism; p. 361-65.
- Auer J, Berent R, Lassnig E, et al. C-reactive Protein and Coronary Artery Disease. Japanese Heart Journal. 2002;43(6):607-19.
- Champe PC, Harvey RA. Lippincott’s Illustrated Reviews : Biochemistry. 2nd ed. Philadelphia: Lippincott; 1994. Chapter 25: Metabolism in the Well-Fed State; p. 281-89.
- Jones EC, Rylands JC, Jardet CL. Regeneration of Pancreatic β-Islet Cells in a Type-II Diabetic. Case Rep Endocrinol. 2018;2018:6147349.
- Tatarinova OV, Kylbanova ES, Neustroeva VN, et al. [Phenomenon of super-longevity in Yakutia]. Adv Gerontol. 2008;21(2):198-203.
- Cha YJ, Kim ES, Koo JS. Amino Acid Transporters and Glutamine Metabolism in Breast Cancer. Int J Mol Sci. 2018;19(3):907.
- S. Food and Drug Administration. Mercury Levels in Commercial Fish and Shellfish (1990-2012).
- https://www.evenbetternow.com/
- Bild W, Stefanescu I, Haulica I, et al. Research concerning the radioprotective and immunostimulating effects of deuterium-depleted water. Rom J Physiol. 1999; 36(3-4):205-18.
- Best and Worst Cold Water Fish to Eat. MD health.
- Boros LG, D’Agostino DP, Katz HE, et al. Submolecular regulation of cell transformation by deuterium depleting water exchange reactions in the tricarboxylic acid substrate cycle. Med Hypotheses. 2016;87:69-74.
- Cerk IK, Wechselberger L, Oberer M. Adipose Triglyceride Lipase Regulation: An Overview. Curr Protein Pept Sci. 2018;19(2):221-33.
- Basolo A, Poma AM, Bonuccelli D, et al. Adipose tissue in COVID-19: detection of SARS-CoV-2 in adipocytes and activation of the interferon-alpha response. J Endocrinol Invest. 2022;45(5):1021-29.
- Zechner R, Kienesberger PC, Haemmerle G, et al. Adipose triglyceride lipase and the lipolytic catabolism of cellular fat stores. J Lipid Res. 2009;50(1):3-21.
- Kintscher U, Foryst-Ludwig A, Haemmerle G, et al. The Role of Adipose Triglyceride Lipase and Cytosolic Lipolysis in Cardiac Function and Heart Failure. Cell Rep Med. 2020;1(1):100001.
- Kolb H. Obese visceral fat tissue inflammation: from protective to detrimental? BMC Med. 2022;20(1):494.
- Eckman MH, Lip GY, Wise RE, et al. Impact of an Atrial Fibrillation Decision Support Tool on thromboprophylaxis for atrial fibrillation. Am Heart J. 2016;176:17-27.
- https://www.mdcalc.com/calc/801/cha2ds2-vasc-score-atrial-fibrillation-stroke-risk
- Cofresí RU, Bartholow BD, Fromme K. Female drinkers are more sensitive than male drinkers to alcohol-induced heart rate increase. Exp Clin Psychopharmacol. 2020;28(5):540-52.
- Zhenyu Z, Caihong S, Zhongming L, et al. Effects of deuterium-depleted liquor on glucose metabolism, islet cells and their functions in diabetic rats. J. Shanghai Jiaotong Univ. 2010;30(10):1204-07.
- Garrett-Bakelman FE, Darshi M, Green SJ, et al. The NASA Twins Study: A multidimensional analysis of a year-long human spaceflight. Science. 2019;364(6436):eaau8650.
- Căluşer I, Suciu I, Cäluşer L. Influence of deuterium on the spleen immune system. Morphol Embryol (Bucur). 1979;25(4):365-68.
- Nielsen J. Bioactive metabolites: The double-edged sword in your food. Cell. 2022;185(24):4469-71.
