Collection and morphology
During a survey, to study the diversity of aquatic fungi, decaying woody specimens were collected from eastern Thailand in December 2018, following the procedures described in Kurniawati et al. [15]. Specimens were placed in ziploc plastic bags with sterile wet cotton and taken to the laboratory. Specimens were morphologically observed using a Nikon (Model SMZ-171, Japan) dissecting microscope to locate fruiting bodies. Photomicrographs were made with a Nikon (Model ECLIPSE Ni, Japan) compound microscope with a Cannon EOS 600D camera. Tarosoft (R) Image Frame Work program was used to measure the fungal structures and Adobe Photoshop CS6 Extended version 12.0 software (Adobe Inc., USA) was used to make the photo plates.
Isolation
Isolations were made from single conidium on potato dextrose agar (PDA) following the methods of Chomnunti et al. [16], and then incubated overnight in an incubator (25°C). Single germinating conidia were transferred to new PDA plate. Cultures were examined and colony characteristics were recorded every seven days.
Dry specimens were deposited in the herbarium of Mae Fah Luang University (MFLU), Chiang Rai, Thailand. Living cultures were deposited in Mae Fah Luang University Culture Collection (MFLUCC) and Kunming Institute of Botany Culture Collection (KUMCC).
DNA extraction, PCR amplification and sequencing
Total DNA was extracted from cultures grown on PDA for 30 days at 25°C using a Biospin Fungus Genomic DNA Extraction Kit (Bioer Technology Co., Ltd., Hangzhou, P.R. China) according to the manufacturer’s instructions. Two molecular markers were used in this study for molecular identification: the internal transcribed spacer (ITS) and the large subunit ribosomal RNA gene (LSU) and their respective primer pairs used were ITS5/ITS4 [17] and LROR/LR5 [18]. The amplifications were carried out in a 25 μL reaction volume containing 9.5 μL ddH2O, 12.5 μL 2 × PCR Master Mix, 1 μL of DNA template, 1 μL of each primer (10 μM). The PCR thermal cycles for amplification of the gene regions followed according to Su et al. [19]. The PCR products were examined on 1.0 % agarose gel stained with Gel Red ethidium bromide. Sequencing reactions were conducted by TSINGKE Biological Engineering Technology and Services Co., Kunming, P.R. China.
We provide the similarity of ITS sequences of each taxon in BLASTn search as a percentage or compared ITS sequences between our new isolates and type strains. Classification of each taxon is also provided based on Wijayawardene et al. [20].
Taxonomy and discussion
- Conlarium aquaticum Dong, H. Zhang & K.D. Hyde, Fungal Diversity 85: 92 (2017) (Figure 1)
Index Fungorum Registration Identifier 553759
Classification: Annulatascaceae, Annulatascales, Diaporthomycetidae, Sordariomycetes
Saprobic on decaying submerged wood in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetes, Colonies sporodochial, broadly and irregularly punctiform, gregarious or scattered, raised, dark brown to black, velvety. Mycelium mostly immersed in natural substratum, consisting of branched, septate, thin-walled, smooth, pale brown to brown hyphae. Conidiophores absent or reduced to conidiogenous cells. Conidiogenous cells up to 18 μm long, monoblastic, holoblastic, integrated, determinate, cylindrical, hyaline to pale brown, smooth. Conidia acrogenous, solitary, dry, subglobose, ellipsoidal, oblong or irregular, brown, clathrate, muriform, 6–12-transversely septate, 4–10-longitudinally septate, slightly constricted at septa, smooth, thin-walled, 47–72 × 25–54 μm ( = 61.5 × 36 μm, n = 30).
Culture characteristics: On PDA, colonies circular, reaching 10 mm in 30 days at 25°C in the condition of 12 h dark and 12 h light; gray to dark brown from above, dark brown from below, umbonate, rough, wrinkled, dry, edge undulate. Conidiophores macronematous, branched, sometimes reduced to a single conidiogenous cell, hyaline. Conidiogenous cells holoblastic, monoblastic, cylindrical, smooth-walled. Conidia acrogenous, solitary, dry, subglobose, ellipsoidal, oblong or irregular, muriform, smooth, thin-walled, hyaline when young, olivaceous at maturity, 42–65 × 25–56 μm ( = 53 × 34 μm, n = 20).
Material examined: Thailand, Mukdahan Province, small river of Nong Bo Na Kae, 16°84’96’’N, 104°33’12’’E, on dead submerged decaying wood of unidentified plants, 13 December 2018, Hao Yang, t35 (MFLU 19–0545), living in culture = KUMCC 19–0209.
Sequences data: ITS: MN513038, LSU: MN511740.
Notes: The genus Conlarium was established to accommodate the holomorphic species C. dupliciascosporum [21]. Zhang et al. [14] introduced the second species C. aquaticum from freshwater. A comparison of nucleotide sequences between our isolate and the type strain of C. aquaticum showed only 4 bp differences without gaps in ITS gene region (https://blast.ncbi.nlm.nih.gov/Blast.cgi), which means they are the same species according to Jeewon et al. [22]. In addition, they have similar morphology including conidial size. Thus, we confirm the identity of our isolate as C. aquaticum. The cultural characteristics of C. aquaticum are provided in this paper, which shows different conidial colour (brown on natural substrate vs. hyaline to olivaceous from culture).
Known distribution: Thailand
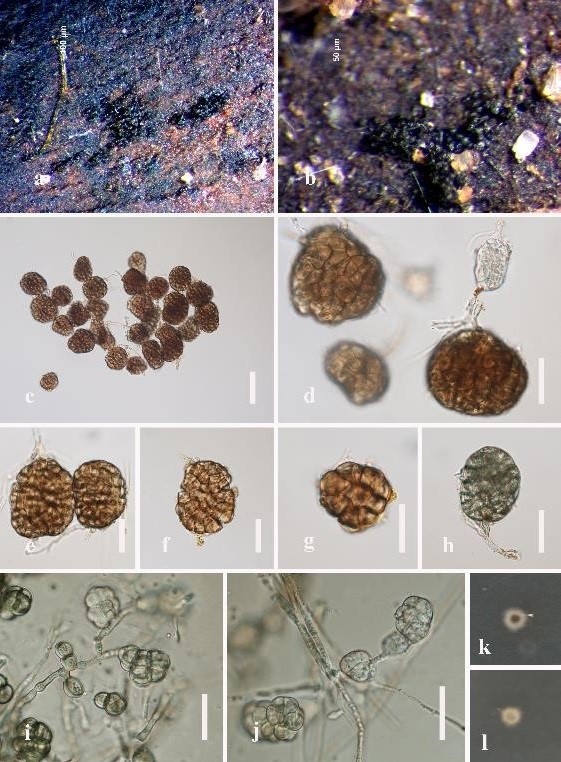
Figure 1: Conlarium aquaticum (MFLU 19–0545). (a), (b) Sporodochia on submerged wood. (c), (d) Conidia attach to conidiogenous cells. (e–h) Conidia. (i), (j) Conidia from PDA. (k), (l) Culture on PDA, k obverse and l reverse. Scale bars: c = 50 μm, d–j = 30 μm.
- Dictyocheirospora nabanheensis Tibpromma & K.D. Hyde, Fungal Diversity 93: 10 (2018) (Figure 2)
Index Fungorum Registration Identifier 554474
Classification: Dictyosporiaceae, Pleosporales, Pleosporomycetidae, Dothideomycetes
Saprobic on decaying wood in freshwater habitats. Sexual morph: undetermined. Asexual morph: hyphomycetous, Colonies sporodochial, scattered or in small groups, dark brown to black. Mycelium mostly immersed. Conidiophores micronematous, reduced to conidiogenous cell. Conidiogenous cells holoblastic, cylindrical, hyaline to pale brown, partly remaining attached to the conidia, 9.5–14 × 4–7 μm. Conidia solitary, oval to ellipsoid, cheiroid, 43.5–55.5 × 20–29.5 μm ( = 50.0 × 24.5 μm, n = 40), consisting of 40–48 cells, with a basal connecting cell, pale brown to yellow-brown, smooth-walled, individual cells discoid, 28–37.5 × 5–6 μm, arranged in 6 compact rows, with 6–10 cells per row; with 1–2 rounded to cylindrical appendages, 5–10 × 4–5 μm, arising from near middle of conidial rows, hyaline.
Cultural characteristics: Conidia germinated on PDA within 24 h. Germ tubes produced from the base cell. Colonies on PDA reaching 18–20 diameter in 18 days at 20–25°C in the condition of 12 h dark and 12 h light, with dense, yellow to brown mycelium on the surface; reverse brown to dark brown with white margin.
Material examined: Thailand, Mukdahan Province, small river of Sakon Nakhom, 16°53’30’’N, 104°11’25’’E, on dead submerged decaying wood of unidentified plants, 13 December 2018, Hao Yang, t24 (MFLU 19–0538), living culture = KUMCC 19–0208.
Sequences data: ITS: MN513035, LSU: MN511737.
Notes: Dictyocheirospora nabanheensis was introduced by Tibpromma et al. [23] and characterised by its cheiroid conidia consisting of 40–48 cells, 6–10 cells in each row and 1–2 rounded to cylindrical appendages. In a BLASTn search on NCBI GenBank, the closest match of our isolate in ITS region is 100% identical to the strain of D. nabanheensis (MK347756); thus, it is identified as D. nabanheensis.
Known distribution: Thailand
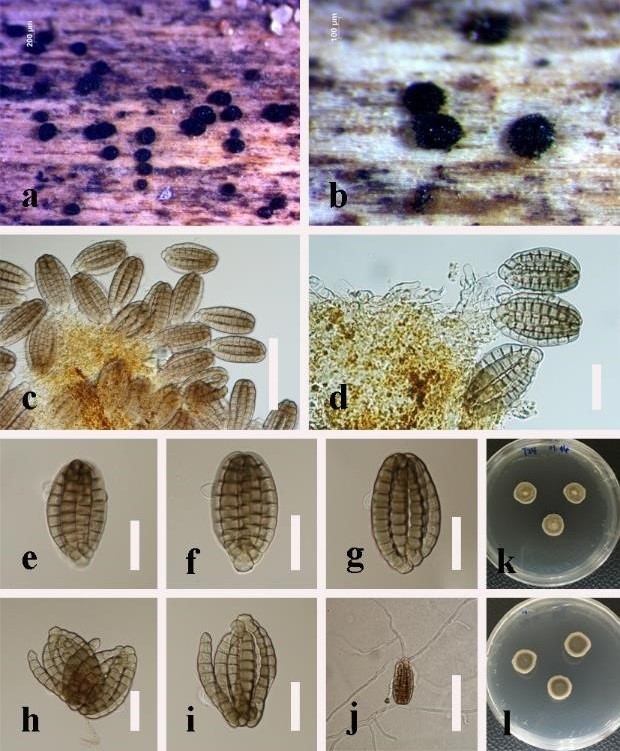
Figure 2: Dictyocheirospora nabanheensis (MFLU 19–0538). (a), (b) Colonies on substrate. (c) Squash of colonies. (d) Conidiogenous cells.
(e–i) Conidia. (j) Germinating conidium on PDA. (k), (l) Culture on PDA, k obverse and l reverse. Scale bars: c, j = 50 μm, d–i = 20 μm.
- Distoseptispora rayongensis Yang & K.D. Hyde, Fungal Diversity (2019), in press. (Figure 3)
Classification: Distoseptisporaceae, Pleosporales, Pleosporomycetidae, Dothideomycetes
Saprobic on decaying wood submerged in freshwater. Sexual morph: undetermined. Asexual morph: hyphomycetous, Colonies on the substratum superficial, effuse, hairy or velvety, black. Mycelium mostly immersed, consisting of branched, septate, smooth, hyaline hyphae. Conidiophores macronematous, mononematous, solitary, unbranched, erect, cylindrical, straight or slightly flexuous, 2–4-septate, brown, smooth, 62.5–73 × 4–5.5 µm ( = 68 × 5 µm, n = 10). Conidiogenous cells monoblastic, integrated, determinate, terminal, cylindrical, brown. Conidia acrogenous, obclavate or obspathulate, rostrate, mostly 13–16-euseptate, rarely 16–24-septate, pale brown or pale olivaceous, becoming paler or hyaline towards the apex, truncate at the base, rounded at the apex, guttulate, smooth, thick-walled, 85–163(–265) × 10–14.5 µm ( = 107 × 12.5 µm, n = 20), with a darkened scar at the base, sometimes with percurrent proliferation and forming another conidium from the conidial apex.
Culture characteristics: Conidia germinated on PDA within 24 h. Germ tubes produced from the conidial top. Colonies on PDA reaching 16 mm diameter after 15 days at 25°C in the condition of 12 h dark and 12 h light, circular, aerial mycelium dense, brown; reverse dark brown with entire white margin.
Material examined: Thailand, Mukdahan Province, small river of Nong Bo Na Kae, 16°84’96’’N, 104°33’12’’E, on dead submerged decaying wood of unidentified plants, 13 December 2018, Hao Yang, t33 (MFLU 19–0543), living culture = KMUCC19–0334.
Sequences data: ITS: MN513037, LSU: MN511739.
Notes: Distoseptispora rayongensis was introduced by Hyde et al. [24] which was found in Rayong Province, Thailand. Our isolate was collected in Mukdahan Province of Thailand and its morphological characteristics, such as cylindrical, unbranched, septate conidiophores and distoseptate conidia, were identical to D. rayongensis. The nucleotide sequences of our isolate were 100% identical to the type strain of D. rayongensis in ITS gene region, therefore, we identified the strain as D. rayongensis.
Known distribution: Thailand

Figure 3: Distoseptispora rayongensis (MFLU 19–0543). (a), (b) Colonies on substrate. (c), (d) Conidiophores with conidia. (e) Conidiophore. (f) Conidiogenous cell. (g–l) Conidia. (m) Germinating conidium. (n), (o) Colony on PDA, n obverse and o reverse. Scale bars: c, l–m = 50 μm, d = 30 μm, e–f = 10 μm, g–k = 20 μm.
- Gliocladiopsis pseudotenuis Lombard & Crous, Persoonia 28: 31 (2012) (Figure 4)
Index Fungorum Registration Identifier 564403
Classification: Nectriaceae, Hypocreales, Hypocreomycetidae, Sordariomycetes
Saprobic on decaying wood in freshwater habitats. Sexual morph: undetermined. Asexual morph: hyphomycetous, Colonies on natural substrata effuse, scattered, white, upper part covered with distinct bright white mass of conidia. Mycelium partly immersed, partly superficial. Conidiophores synnematous, micronematous, penicillate, septate, smooth, hyaline, 61–130 × 3–6 μm, without stipe extensions and terminal vesicles. Conidiogenous cells polyblastic, terminal, phialides doliiform to cymbiform to cylindrical, hyaline, 10–17 × 2–3 μm. Conidia cylindrical, 18–21 × 3–4 μm ( = 19 × 3 μm, n = 20), hyaline, smooth with rounded ends, straight, with a single median septum.
Culture characteristics: Conidia were germinated on PDA within 24 h. Colonies fast-growing, attaining 40 mm diameter in 13 days at 20–25°C in the condition of 12 h dark and 12 h light, floccose, rounded, white at first, brown at the center, becoming pale yellow when aged with dense mycelium on the surface, with undulate margin, colonies reverse yellow.
Material examined: Thailand, Mukdahan Province, small river of Nong Bo Na Kae, 16°84’96’’N, 104°33’12’’E, on dead submerged decaying wood of unidentified plants, 13 December 2018, Hao Yang, t37 (MFLU 19–0547), living culture = MFLUCC 19–0337.
Sequences data: ITS: MN513039, LSU: MN511741.
Notes: Gliocladiopsis pseudotenuis was introduced to accommodate G. tenuis (Bugnic.) Crous & M.J. Wingf. [25]. Morphologically, our isolate shares the similar characters to that of G. pseudotenuise except the slightly larger conidia ( = 19 × 3 µm vs. 17× 2 µm). A comparison of nucleotide sequences between our isolate and the holotype of G. pseudotenuis showed a difference of only 3 bases in ITS gene regions, that means they are almost 100% identical, therefore, we validated our strain as G. pseudotenuis.
Known distribution: Hong Kong, Thailand
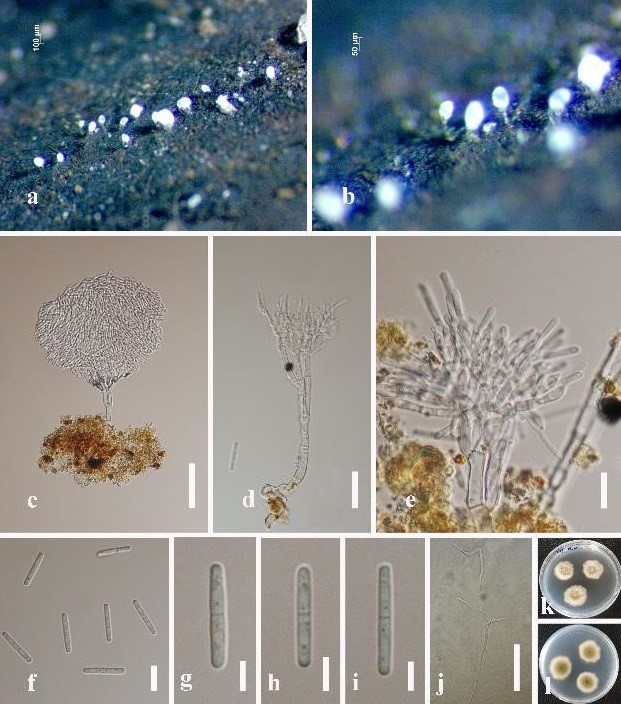
Figure 4: Gliocladiopsis pseudotenuis (MFLU 19–0547). (a), (b) Colonies on substrate. (c), (d) Conidiophores with conidia. (e) Conidiogenous cells. (f–i) Conidia. (j) Germinating conidium. (k), (l) Culture on PDA, k obverse and l reverse. Scale bars: c = 50 μm, d, e = 20μm, f = 10 μm, g–j = 5 μm.
- Spadicoides cordanoides Goh & K.D. Hyde, Mycologia 88: 1023 (1997) (Figure 5)
Index Fungorum Registration Identifier 436825
Classification: Helminthosphaeriaceae, Sordariales, Sordariomycetidae, Sordariomycetes
Saprobic on submerged decaying wood. Sexual morph: undetermined. Asexual morph: hyphomycetous, Colonies on the substratum superficial, effuse, hairy, dark brown to black. Mycelium partly immersed, partly superficial, composed of pale brown, septate, smooth, branched hyphae. Conidiophores macronematous, mononematous, straight or slightly flexuous, solitary or in small groups, thick-wall, smooth, unbranched, multi-septate, pale brown to brown, paler towards the apex, 224–565 × 4–6 μm ( = 424 × 5 μm, n = 20), swollen or nodulose at the apex. Conidiogenous cells polytretic, integrated, terminal and intercalary, determinate. Conidia 1-septate, darkly banded at the septa, acropleurogenous, solitary, versicolorous, thick-walled, predominately 2-celled and obpyriform or ovoid, 15–20 × 9–13 μm ( = 17.5 × 11 μm, n = 40), base cell smooth, subhyaline to pale brown, 4–5 μm long, 5–6 μm wide, more or less obtrapezoidal or subcylindrical, with a truncate base, apical cell enlarged, subglobose, 8–12 μm long, 9–12.5 μm wide, dark brown or almost black, verrucous, with guttule when young.
Material examined: Thailand, Mukdahan Province, Nong Bo Na Kae, 16°84’96’’N, 104°33’12’’E, on dead submerged decaying wood of unidentified plants, 13 December 2018, Hao Yang, t31 (IFRD 500–001).
Notes: Spadicoides cordanoides was introduced by Goh et al. [26]. They also provided a key to included 20 different species of Spadicoides. Subsequently, Ma et al. [27] updated the key of Spadicoides that included 40 species. Based on the distinguishing characteristics of the present strain in comparison to that of the 40 known species, we identify our isolate as S. cordanoides.
Known distribution: Australia, Thailand

Figure 5: Spacicoides cordanoides (IFRD 500–001). (a), (b) Colonies on substrate. (c–f) Conidiophores and Conidia. (g–i) Conidiogenous cells and conidia. (j–o) Conidia. Scale bars: c = 100 μm, d–f = 50 μm, g–h, k–o = 10 μm, i–j = 20 μm.
- Sporoschisma longicatenatum Jiao Yang, Jian K. Liu & K.D. Hyde. Phytotaxa 289: 152 (2016) (Figure 6)
Index Fungorum Registration Identifier 552182
Classification: Chaetosphaeriaceae, Chaetosphaeriales, Sordariomycetidae, Sordariomycetes
Saprobic on submerged decaying wood. Sexual morph: undetermined. Asexual morph: hyphomycetous, Colonies on the substratum superficial, effuse, hairy, gregarious, black, with long chains of conidia. Mycelium immersed, composed of dark brown hyphae. Setae scattered or in groups mixed with conidiophores, capitate, usually with hyaline, mucilaginous substances at the swollen apex, smooth-walled, pale brown, becoming paler towards the apex, straight or flexuous, 2−3-septate, 113−168 × 4.5–6 μm, 6–8.5 μm wide at the swollen apex. Conidiophores macronematous, mononematous, smooth, dark brown to black, straight or slightly flexuous, solitary or in small groups of 2−3, each composed of a bulbous base, a cylindrical stipe and a swollen venter with a long cylindrical neck, erect, 259–316 μm long, 7.5–12 μm wide below venter and 13–15.5 μm wide above, 16–21 μm wide at venter. Conidiogenous cells monophialidic, percurrent, integrated, terminal, determinate, brown, lageniform, frayed at the apex, with serrate, flared margin at free end. Conidia formed in chains, cylindrical to doliiform, 31–47 × 9–12.5 μm ( = 41 × 10 μm, n = 30), 5-euseptate, hyaline when young, olivaceous to brown at maturity, with conspicuously darkened septa, occasionally constricted at septa, centre cells brown, end cells paler coloured and shorter than the central four cells.
Culture characteristics: Conidia germinating on PDA within 24 h and germ tubes produced from both ends. Colonies on PDA attaining 15 mm diameter in 15 days at 20–25°C in the condition of 12 h dark and 12 h light, with fluffy, dense, white mycelium on the surface, colonies reverse dark brown at the centre, with white undulate margin.
Material examined: Thailand, Mukdahan Province, Nong Bo Na Kae, 16°84’96’’N, 104°33’12’’E, small river, on dead submerged decaying wood of unidentified plants, 13 December 2018, Hao Yang, T29 (MFLU 19–0540), living culture = MFLUCC 19–0331.
Sequences data: ITS: MN513036, LSU: MN511738.
Notes: Sporoschisma longicatenatum was introduced by Yang et al. [28]. Even though, our isolate shares similar morphological characters with S. hemipsilum (Berk. & Broome) Zelski, A.N. Mill. and Shearer and S. longicatenatum. The closest match of ITS sequence of our strain is 100% identical to S. longicatenatum strain MFLUCC 16–0180 (KX505871) in the BLASTn search on NCBI GenBank, thus, we identified our strain as S. longicatenatum.

Figure 6: Sporoschisma longicatenatum (MFLU 19-0540). (a), (b) Colonies on submerged wood. (c), (d) Conidiophores and setae. (e) Portion of phialide producing conidium. (f) Conidiophore with conidia. (g–k) Conidia. (l) Germinating conidium on PDA. (m), (n) Culture on PDA, m from front, n from reverse. Scale bars: c, d = 100 μm, e–g = 50 μm, h–l = 20 μm.
