Mesole SB1,3,4, Ibegbu AO2, Musa SA3,4, Bauchi ZM3,4, Agbon AN3,4, Okpanachi OA5* and Ivang A6
1Department of Clinical Medicine, School of Medicine and Health Sciences, Eden University, Lusaka, Zambia
2Department of Anatomy, Faculty of Basic Medical Sciences, Alex Ekueme University Ndufu Alike Ikwo, Nigeria
3Department of Anatomy, Faculty of Basic Medical Sciences, Ahmadu Bello University, Zaria, Nigeria
4Neuroanatomy and Neuroscience Unit, Department of Human Anatomy, Ahmadu Bello University, Zaria, Nigeria
5Department of Physiology, Faculty of Biomedical Sciences, Kampala International University, Uganda
6Clinical Anatomy Unit, Department of Clinical Biology, College of Medicine and Health Sciences, University of Rwanda, Rwanda
*Correspondence: Omachonu Alfred Okpanachi, Department of Physiology, Faculty of Biomedical Sciences, Kampala International University, Uganda
Received on 17 February 2021; Accepted on 02 June 2021; Published on 09 June 2021
Copyright © 2021 Mesole SB, et al. This is an open access article and is distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Abstract
The metal aluminium (Al) is a well-documented neurotoxin and has been known to exacerbate excitotoxicity within the human nervous system. This present research is aimed and intended at assessing the neuroprotective effect of eugenol (EG) against the toxic effects of aluminium chloride (AlCl3) in vivo using Wistar rats as a study model. 20 adult Wistar rats were randomly assorted or divided into 4 groups, and each group contains 5 animals. This study was conducted for a duration of 21 days. On day 22 (24 h after the last administration), rats were compassionately sacrificed with 0.8 mg/kg ketamine as anaesthesia. Thereafter brain tissue was removed for immunohistochemistry and various biochemical estimations which included antioxidant enzyme superoxide dismutase (SOD), glutathione peroxidase (GPx), intra-mitochondrial accumulation of 8-hydroxy-2-deoxyguanosine (8-OHdG), pro/anti-apoptotic proteins Bax, Bcl-2 and caspase-3, endogenous enzymatic activity of acetylcholinesterase (AChE), immunohistochemistry of acetylcholine using acetyl-CoA acetyltransferase-1 (ACAT-1) within the cornu Ammonis-1 (CA1) region of the hippocampus and cognitive ability using novel object recognition test (NOR). Result obtained revealed a significant (p < 0.05) increase in the activity of antioxidant enzymes, a reduction in pro-apoptotic protein levels upon oral administration of EG. The results obtained show EG as a promising prospect with regards to neurotoxicity as a result of aluminium chloride (AC) toxicity.
Keywords
superoxide dismutase, glutathione peroxidase, caspase-3, acetylcholinesterase, novel object recognition test, hippocampus, immunohistochemistry, eugenol
Abbreviations
Al: aluminium; SOD: superoxide dismutase; GPx: glutathione peroxidase; 8-OHdG: 8-hydroxy-2-deoxyguanosine; AChE: acetylcholinesterase; ACAT-1: acetyl-CoA acetyltransferase-1; NOR: novel object recognition test; CTRL: Control; AC: aluminium chloride; EG: eugenol; ROS: reactive oxygen species; mtDNA: mitochondrial DNA; PBS: phosphate buffered saline; SEM: standard error of mean
Introduction
Aluminium (Al) is highly reactive with both carbon and oxygen, which are among the leading elements required for the sustenance of the biological ecosystem. Due to this, the use of bioavailable Al can have a huge implication towards the health of humans and animals. Scientific study reveals that Al is toxic to all forms of life within our ecosystem, and its appearance in terrestrial biochemistry also reveals an invariably injurious effect [1].
The relationship between the neurotoxicity and consequent excitotoxicity of Al neurotoxicity was discussed by Exley [2]. Meanwhile, Al, under certain conditions can act as a pro-oxidant. However, Al has the capacity to react with superoxide radical (O2−) and therefore expedite its destructive ability within a biological system [2].
Daily human use of Al metal in various ways can include but not limited to the manufacture of drinking cans, cookware, aluminium foil, housing materials, and components of electrical devices, airplanes, boats, cars [3, 4].
Various observations from different studies revealed that the formation and generation of superoxide (O2−) is a main determinant and a contributor which promotes excitotoxicity enhanced neuronal death which can occur through the generation of peroxynitrite [5]. The major primary detrimental reaction within the excitotoxic process is the build-up of peroxynitrite, which is a by-product that results from elevated and accumulated levels of superoxide (O2−) and its reaction with nitric oxide (NO) which is also a by-product of excitotoxicity [2]. Excitotoxicity can also be enhanced by Al among cells of the central nervous system (CNS) via the induction of the apoptotic cascade within astrocytes. These astrocytes are the major target of accumulation of Al among various neuronal cell types [6]. Astrocytes are a major source of glutathione and they (glutathione) are required for the proper functioning of neurons and glutathione levels have been shown to be depleted by Al [7].
Eugenol (EG) as a nutraceutical, has been touted to possess a very promising anti-cancer property and as a result, this has resulted in increased attention on EG over the years. Anti-cancer drugs produced synthetically have been known to have toxic side effects such as hair loss, nausea and vomiting, fatigue and constipation, and as a result, there is damage to neuronal cells within the nervous system. EG hence has a preferred use and potential for preventing and treatment of some type of cancers. EG prevents apoptosis by adequately controlling the activity levels of COX-2, Bcl-2, and IL-1β, which will result in reduction of inflammation and consequently preventing death of various neurons in the nervous system [8].
This present research is aimed at investigating the efficacy of EG in protecting neurons of the nervous system from toxic abuse as a result of oral administration of aluminium chloride (AC). This research explores the ameliorative potential and the preventive approach to diminish or reduce the deleterious effects of Al on the brain.
Materials and Methods
Animals
20 adults male Wistar rats with an average weight of 140–160 g were obtained from the animal house of Ahmadu Bello University, Zaria, Nigeria, with unlimited or free access to food and water. Experiments were carried out in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 80-23), revised 1996, and all measures were put in place to significantly reduce animal suffering [9]. Wistar rats were randomly divided into 4 groups, each group containing 5 animals each as shown below (Table 1).
Control (CTRL) 0.6 ml/kg of distilled water, 100 mg/kg of aluminium chloride (AC), 300 mg/kg of eugenol (EG) + 100 mg/kg of aluminium chloride (AC) and 300 mg/kg eugenol (EG) only.
| Group | Dose |
| Control (CTRL) | 0.6 ml/kg (distilled water) |
| Aluminium chloride (AC) | 100 mg/kg |
| Eugenol + aluminium chloride (EG + AC) | 300 mg/kg + 100 mg/kg |
| Eugenol (EG) | 300 mg/kg |
Table 1: Animal grouping and dosage.
On day 22 (24 h after the last administration), rats were humanely sacrificed with 0.8 mg/kg ketamine as an anaesthetic agent. Rat brains were removed carefully, adhering structures were also removed and washed twice (2x) in 50 mM Tris-HCl (pH 7.4). Brain tissue removed was weighed with a sensitive digital weighing balance and consequently homogenized to obtain a 10% (w/v) homogenate in ice-cold medium. The resultant homogenate was centrifuged at the rate of 1200 g for a period of 10 min at a temperature of 4°C. Supernatants that was obtained was used for various enzymatic and protein estimations. The overall protein content of the resultant homogenate was assayed by the method of Al-Olayan et al. [10], which was reported by Mæhre et al. [11] adopting bovine serum albumin as a standard during the assay.
Oxidative stress markers
A biological system combats oxidative stress with the use of a different enzymatic process that uses various antioxidants which includes but not limited to superoxide dismutase (SOD) and as a result SOD enzyme level or activity was assayed or measured using the correct Rat SOD ELISA kit which was supplied by WKEA Med Supplies Corp, China and the kit was manufactured by FineTest China. These kits were used in line with instruction from the manufacturer. The procedure used in this study was based on the method of Okey et al. [12] and reported by Kumar et al. [13] and Mesole et al. [14]. Glutathione peroxidase (GPx) which is also an antioxidant enzyme, was assayed using the appropriate Rat GPx ELISA kit which was supplied by WKEA Med Supplies Corp, China and manufactured by FineTest China. This kit was used in strict compliance with instructions from the manufacturer. The procedure adopted was based on the method of Odinga et al. [15]. Consequently, the enzymatic levels of GPx and SOD were expressed as µmol/ml.
Generation of reactive oxygen species (ROS) can also result in damage to mitochondrial DNA (mtDNA). This damage is quantified by assaying the tissue levels of 8-hydroxy-2-deoxyguanosine (8-OHdG) using the appropriate Rat 8-OHdG ELISA kit which was supplied by WKEA Med Supplies Corp, China and manufactured by FineTest China. Isolation of mtDNA was carried out by the method which was described by Odinga et al. [15] as reported by Mesole et al. [14] and Rui et al. [16].
Determination of apoptotic markers
Homogenates of brain tissues are made with lysis buffer and were evaluated using a Rat Colorimetric Caspase-3 Assay kit supplied by WKEA Med Supplies Corp, China and manufactured by FineTest China. These kits were used with strict compliance to the manufacturer’s instruction. Anti-apoptotic and pro-apoptotic protein (Bcl-2 and Bax) levels was assayed or analyzed in brain tissue lysates using appropriate Rat ELISA kit supplied by WKEA Med Supplies Corp, China and manufactured by FineTest China. Assay of these proteins was carried out according to the manufacturer’s instruction and results obtained were expressed as ng/mg.
Endogenous enzyme (acetylcholinesterase)
The tissue activity or levels of the enzyme acetylcholinesterase (AChE) was evaluated using Rat ELISA Acetylcholinesterase Activity Assay kit (Sigma-Aldrich, USA). This kit was used in strict compliance to manufacturer’s instruction. The procedure adopted was based on the method used by El-Marasy et al. [17] and reported by Zhao et al. [18] and results obtained were expressed as µmol/ml.
Immunohistochemical studies
Immunohistological sections of Wistar rat brain region were incubated with acetyl-CoA acetyltransferase-1 (ACAT-1) antibody, then counter-stained with routine haematoxylin and eosin (H&E) and examined under the light microscope. Staining protocol is highlighted below.
Histological sections were sectioned at a thickness of 4 microns using a microtome. These sections were placed on electrostatically charged slides and these slides were baked at 70°C for 10 min and the slides were taken to distilled water. After removed from distilled water, slides were blocked in hydrogen peroxide for a duration of 5 min and were washed in distilled water again. Afterwards, the slides were retrieved in citrate buffer pH 6.0 for a period of time duration of 20 min at a temperature of 95°C and the slides were washed in distilled water and then the slides were covered with phosphate buffered saline (PBS) pH 7.4 and slides were incubated with ACAT antibody (1/100) at RT for a duration of 90 min. Post-incubation, slides containing tissues were washed in PBS at pH 7.4 and then incubated in Mouse + Rabbit HRP (DB Biotech, Slovakia), for a period of time duration of 30 mins.
Slides were washed in PBS for a duration of 10 min and subsequently washed slides were incubated in DAB + substrate mixture (DB Biotech, Slovakia) for a duration of 5 mins, rewashed using distilled water again and counter stained with haematoxylin for a duration of 1 min. After counterstaining, slides were washed in distilled water, dehydrated (removing any residual fluid), cleared and mounted in DPX.
Novel object recognition test (NOR)
Various behavioral tests are used to assess various behavioral end points. End points that can be assessed include anxiety, learning, and memory. Commonly used behavioral test for rats is the object recognition test to assess recognition memory. This test is made up of two sessions. In both sessions, a rat is placed in a box and presented with two (A and B) objects that look alike (similar object) during the first session and in the second session one of the objects is replaced by a new (novel) object (C). The time duration spent exploring the new object is a measure for an index of recognition memory [19].
Procedure
The method adopted for this study was described by de Almeida et al. [20] and Labban et al. [21]. All rats used were habituated to the set-up for a period of 5 min each. The rats were then carried to the test room in their cages and the tests were carried out individually. 5 min each were allotted to each rat and then returned back to their cages. The apparatus was cleaned after each test with 70% ethyl alcohol and allowed to dry between trials.
Statistical analysis
Results obtained from this research were analyzed with just another statistical program (JASP). These results were expressed as the mean ± standard error of mean (SEM) and the resultant significant difference obtained among the obtained value of the mean of the groups was determined with one-way analysis of variance (ANOVA) with repeated measures using Turkey’s post hoc test for significance. Paired sample t-test was employed for the comparison of the means. Values were considered significant when p ≤0.05.
Results
The figure reveals the effect of EG on tissue (brain) activity/levels of SOD after oral intake of AC (Figure 1). The result obtained revealed a significant (p < 0.05) reduction in tissue (brain) activity of SOD among the group that received 100 mg/kg AC when comparisons were made to the CTRL group. However, co-administration of EG + AC resulted in a significant (p < 0.05) increase in brain tissue activity/levels of SOD when compared to AC group. Within the group administered 300 mg/kg EG only resulted in significant (p < 0.05) increase in tissue activity of SOD when compared to the CTRL and AC group respectively.
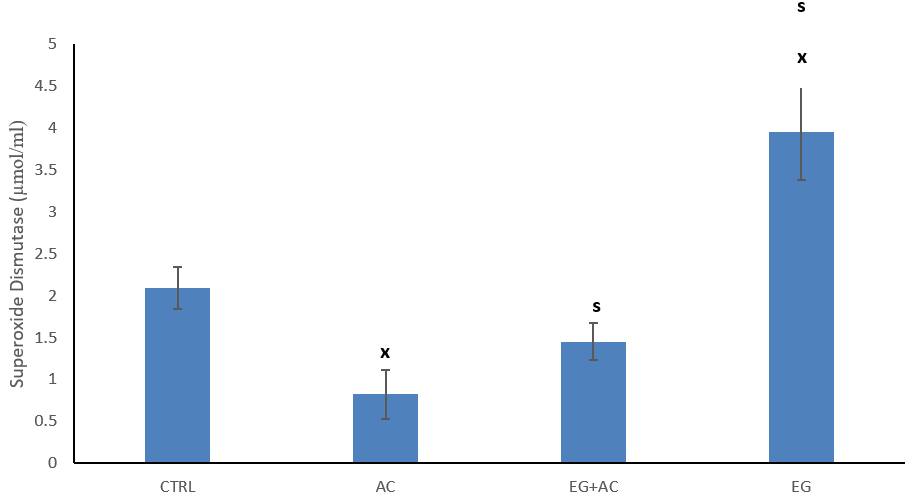
Figure 1: Effect of eugenol (EG) on tissue (brain) superoxide dismutase (SOD) concentration following administration of aluminium chloride (AC). n = 5; mean ± SEM one-way ANOVA LSD post hoc test: s = p < 0.05 when compared with aluminium chloride (AC) treated group; x = p < 0.05 when compared with the control (CTRL).
The figure shows the effect of EG on tissue (brain) GPx concentration following administration of AC (Figure 2). The result shows a significant (p < 0.05) reduction in tissue activity of GPx when compared to CTRL. Co-administration of EG + AC resulted in a significant (p < 0.05) increase in activity of SOD when compared to the AC group. Within the group administered 300 mg/kg EG resulted in a significant (p < 0.05) increase in the activity of SOD when compared to the AC group.
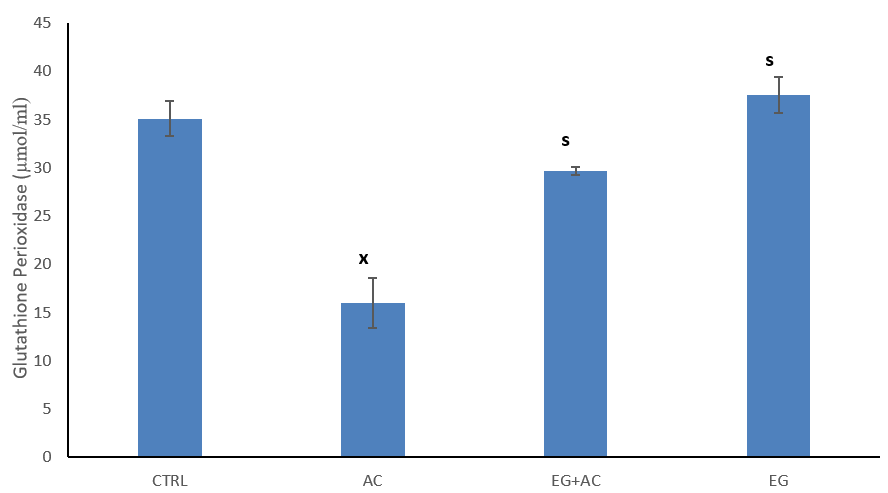
Figure 2: Effect of eugenol (EG) on tissue (brain) glutathione peroxidase (GPx) concentration following administration of aluminium chloride (AC). n = 5; mean ± SEM one-way ANOVA LSD post hoc test: s = p < 0.05 when compared with aluminium chloride (AC) treated group; x = p < 0.05 when compared with the control (CTRL).
The figure shows the effect of EG on intramitochondrial accumulation of 8-OHdG levels following exposure to AC (Figure 3). Oral administration of AC resulted in a significant (p < 0.05) increase in the activity of 8-OHdG when compared to CTRL. Co-administration of EG + AC resulted in a significant (p < 0.05) decrease in tissue levels of 8-OHdG when compared to CTRL. Administration of 300 mg/kg of EG resulted in a significantly (p < 0.05) decreased level of 8-OHdG when compared to AC group.
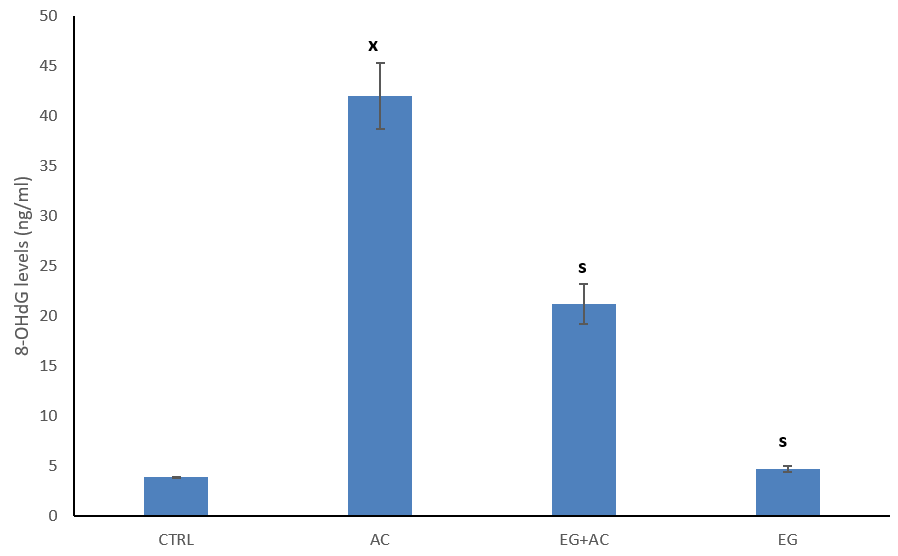
Figure 3: Effect of eugenol (EG) on intramitochondrial accumulation of 8-hydroxy-2-deoxyguanosine (8-OHdG) levels following exposure to aluminium chloride (AC). n = 5; mean ± SEM one-way ANOVA LSD post hoc test: s = p < 0.05 when compared with the aluminium chloride (AC) treated group; x = p < 0.05 when compared with the control (CTRL).
The figure shows the effect of EG on caspase-3 levels in the brain following administration of AC (Figure 4). Oral administration of AC resulted in a significant (p < 0.05) increase in the level of caspase-3 when compared to CTRL group. Co-administration of EG + AC resulted in a significant (p < 0.05) reduction in caspase-3 levels when compared to CTRL but non-significant when compared to AC group. Administration of EG only resulted in a significant (p < 0.05) reduction in the levels of caspase-3 when compared to AC group.
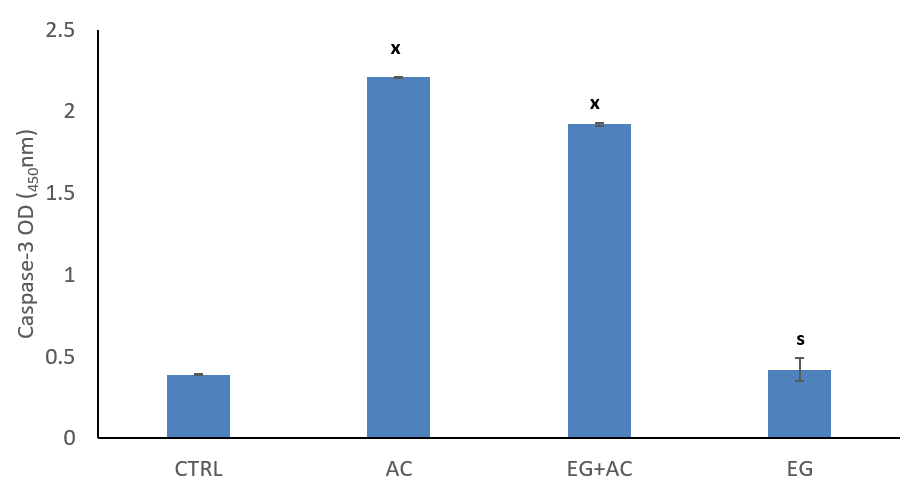
Figure 4: Effect of eugenol (EG) on brain caspase-3 levels following administration of aluminium chloride (AC) in Wistar rats. n = 5; mean ± SEM one-way ANOVA LSD post hoc test: s = p < 0.05 when compared to the aluminium chloride (AC) treated group; x = p < 0.05 when compared to the control (CTRL).
The table shows the effect of EG on pro-apoptotic and anti-apoptotic proteins following administration of AC (Table 2). This result shows a significant (p < 0.05) reduction in the level of anti-apoptotic protein Bcl-2 among the AC treated group when compared to the CTRL. Co-administration of EG and AC resulted in a non-significant (p > 0.05) increase in Bcl-2 when compared to AC group, but a significant (p < 0.05) when compared to CTRL group. Administration of EG only resulted in significant (p < 0.05) change in the Bcl-2 levels when compared to AC and CTRL groups. Administration of AC resulted in a significant (p < 0.05) increase in the levels of pro-apoptotic protein Bax when compared to the CTRL group. Co-administration of EG + AC, however, resulted in a significant change in Bax protein levels (p < 0.05) when compared to AC and CTRL groups respectively. Administration of EG only resulted in changes in Bax protein levels, which is significant (p < 0.05) when compared to the AC group.
| Pro/anti-apoptotic proteins | CTRL | AC | EG + AC | EG |
| Bcl-2 | 17.06 ± 0.62 | 5.12 ± 0.12x | 6.02 ± 0.72y | 13.25 ± 0.21qx |
| Bax | 3.21 ± 0.82 | 19.52 ± 0.60x | 15.21 ± 0.81qy | 4.36 ± 0.37q |
| Bax/Bcl-2 | 0.19 ± 0.01 | 3.81 ± 0.05x | 2.53 ± 0.09qx | 0.32 ± 0.05r |
Table 2: Effect of eugenol (EG) on pro-apoptotic and anti-apoptotic proteins following administration of aluminium chloride (AC) in Wistar rats. n = 5; mean ± SEM one-way ANOVA LSD post hoc test: q, r, s = p < 0.05; when compared with aluminium chloride (AC) treated group; x, y = p < 0.05 when compared to the control (CTRL) group.
The figure shows the effect of EG on brain AChE concentration following the administration of AC (Figure 5). This result showed a significant (p < 0.05) increase in tissue activity of AchE within the AC group when compared to CTRL. Co-administration of EG + AC resulted in a significant (p < 0.05) reduction in brain AchE activity when compared to the AC group. Administration of EG only also resulted in a significant (p < 0.05) reduction in AchE levels when compared to AC group.
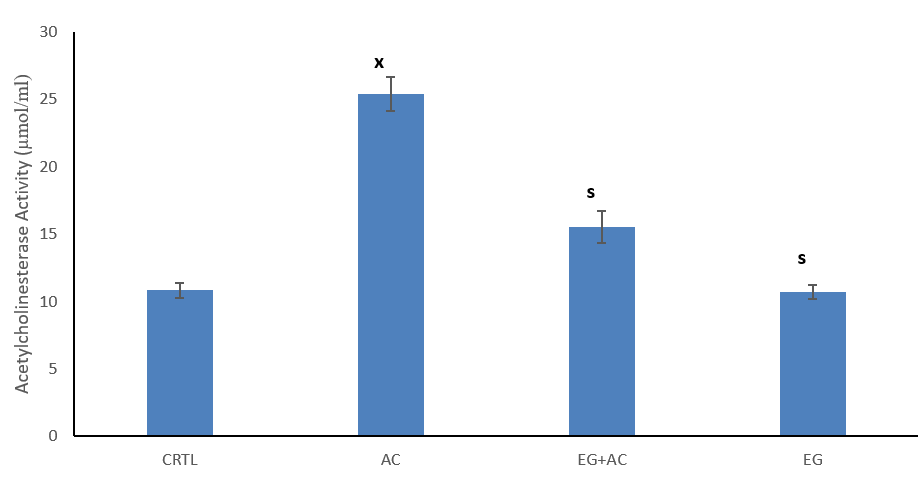
Figure 5: Effect of eugenol (EG) on tissue (brain) acetylcholinesterase (AChE) concentration following administration of aluminium chloride (AC) in Wistar rats. n = 5; mean ± SEM one-way ANOVA LSD post hoc test: s = p < 0.05 when compared with aluminium chloride (AC) treated group; x = p < 0.05 when compared with the control (CTRL); AChE = acetylcholinesterase.
Immunostaining reaction with ACAT-1 revealed immunohistochemical localization of acetylcholine within the neuronal cell body.
- CTRL = administered 0.6 ml/kg distilled water showing normal immunoreactivity.
- AC = administered 100 mg/kg of aluminium chloride showing reduced/diminished immunoreactivity.
- EG + AC = administered 300 mg/kg eugenol + 100 mg/kg aluminium chloride with an improved immunoreactivity when compared to the AC group.
- EG = administered 300 mg/kg eugenol only with an almost normal immunoreactivity when compared to the CTRL.
The figure shows the effect of EG on the cognition of Wistar rats using the novel object recognition test (NOR) following administration of AC (Figure 6). This result shows a decrease in time spent in exploring the novel object from pre-treatment to day 21 within the AC group, however on day 21 this decrease was found to be significant (p < 0.05) when compared to the CTRL on day 21. Co-administration of EG + AC resulted in a non-significant change (p > 0.05) in the time spent on the novel object from pre-treatment to day 21. Administration of EG only resulted in a significant (p > 0.05) increase in time spent on the novel object on day 21 when compared to the AC group.
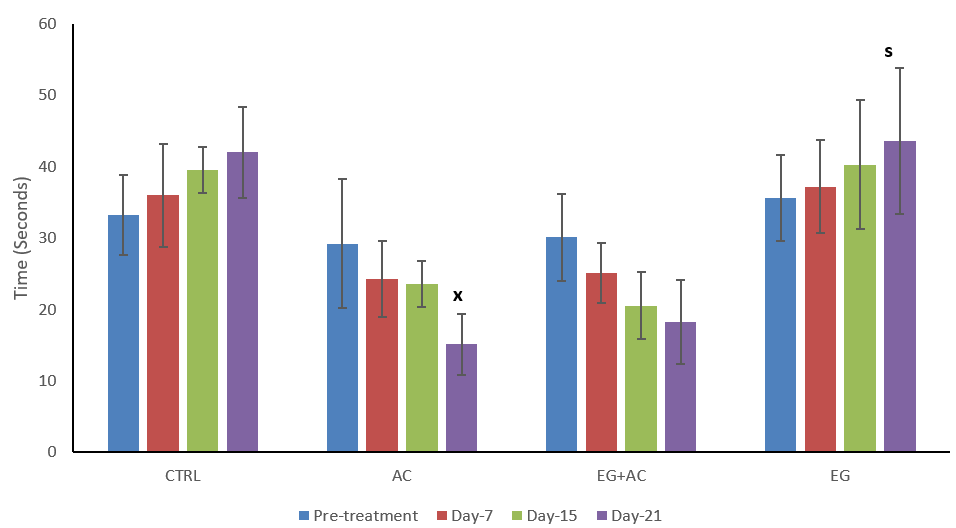
Figure 6: Effect of eugenol (EG) on cognition using the novel object recognition test (NOR) following administration of aluminium chloride (AC). n = 5; mean ± SEM one-way ANOVA with repeated measures and LSD post hoc test: s = p < 0.05 when compared with aluminium chloride (AC) treated group; x = p < 0.05 when compared with the control (CTRL) group on day 21.
Discussion
Different studies carried out showed that excitotoxicity can be greatly increased by reductions within neuronal energy. Disrupting energy metabolism can result to mitochondrial dysfunction and this is one of the mechanisms that is used by Al to amply neuronal excitotoxicity and hence rapidly expedite neuronal cell damage within different brain regions [22–24]. Heavy metals such as mercury and lead are leading pollutants, however this is not limited to heavy metals alone because metals such as Al are also capable of considerable damage due to the fact that these metals cannot be broken down by cells and hence, they are not considered safe for the environment. This makes them play a major role in the aetiology and pathology of various diseases such as neurodegenerative diseases [25].
Various conditions can result in the generation or production of ROS which consequently results in oxidative stress within a biological system. This effect observed can be counteracted by various antioxidants within the biological system [26, 27]. Imbalance between detoxification and ROS production has been associated to different neurodegenerative diseases [28, 29].
In our present research, Wistar rats that received oral administration of different Al salts such as aluminium chloride and aluminium fluoride revealed various indicators of oxidative stress, which was revealed by the reduced level of antioxidants such as SOD and GPx and subsequent reduction in the activities of these antioxidants can lead to generation of ROS [30–32].
Biological systems have the ability to protect themselves against the deleterious effects of activated species in vivo through multiple ways. These include but not limited to free radical scavengers and chain reaction terminators, enzymes such as SOD and GPx systems are one of the few ways this is carried out [33].
The enzyme SOD is part of the enzymatic anti-oxidative defense mechanism that protects against the effects of ROS, and protects various macromolecules, cells and cell membranes from peroxidative damage. The disproportionation of superoxide anion radical into oxygen (O2-) and hydrogen (H+) is catalyzed by SOD [34]. A sensitive index of oxidative damage is the activity of SOD, which scavenges the superoxide anion which forms hydrogen peroxide which results in a reduction in the toxic effects [35].
In this study, SOD tissue activity levels decreased in the aluminium treated group which is in agreement with the study carried out by Garabadu et al. [36] who reported a decrease in the activity of SOD upon administration of AC. Reduction in the activity of antioxidant enzymes such as catalase, SOD, GPx [37]. Administration of EG in this study maintained and elevated the levels of SOD activity. Elevation of SOD activity clearly indicates the antioxidant capacity of EG which is in consistence with the reports of Garabadu et al. [36] and Yuan et al. [ 38].
Oxidative stress and generation of ROS has been a culprit in the pathogenesis of various disease processes and its extent of injury can be related to either an increase or decrease in one or more free radical scavenging enzymes and an example of such enzymes is GPx. GPx possess peroxide activity with a main role to protect the organism from oxidative damage by removing free peroxides in the cell and preventing lipid peroxidation of cellular membranes within a biological system.
In this study, intoxication with AC resulted in a decrease in tissue activity of GPx and co-administration of AC and EG resulted in a significant (p < 0.05) increase in the tissue activity of GPx which confers some level of neuroprotection within the brain of Wistar rat. Results obtained from this study were in accordance with the study conducted by Mesole et al. [14, 39].
The production of free radicals within a biological system can result in damage to proteins and DNA [40]. Mutations within the mtDNA can arise from generation of ROS and this will result in the damage to mtDNA by the virtue of which damage to mtDNA was assessed in this study.
The closeness of mtDNA to site of ROS production makes it very vulnerable to oxidative stress; as a result, the production of ROS is highly toxic to mtDNA [39]. Several agents can result in the discharge of numerous indicators of oxidative stress. The most frequently used indicator of oxidative damage to mtDNA is the production of 8-OHdG [41]. Tissue levels of 8-OHdG within the brain of Wistar rats, which is generated within the mtDNA, are often used as an indicator to determine cellular oxidative damage levels. In the present research, oral administration of AC resulted in significantly increased cellular levels of 8-OHdG. This is in agreement with the study carried out by Mesole et al. [14] and Chiorcea-Paquim et al. [42]. Co-administration of AC with EG, however, resulted in a decrease in cellular levels of 8-OHdG as a result conferring a neuroprotective effect against ROS induced oxidative damage to mtDNA and this is also supported by the research carried out by Mesole et al. [14] in a dose of 150 mg/kg EG.
Analysing the system EG mitigates aluminium induced apoptosis, anti-apoptotic (Bcl-2), pro-apoptotic (Bax), and caspase-3 protein levels were analysed in rat brain homogenates. Production of ROS within a biological system intensifies the permeability of the mitochondrial membrane, which can result in failure within the mitochondrial membrane [41]. Permeability of the mitochondrial membrane is reliant on the level of mitochondrial permeability transition pore, which will facilitate the discharge of cytochrome c from within the inner membrane of the mitochondria into the cytosol located within the cytoplasm [42].
The discharge cytochrome c binds to Apaf-1, these initiates a cascade of events that will culminate in the production of caspase-9, which will in turn lead to the generation of caspase-3 and hence inducing the process of apoptosis [43]. In this study, it was observed that oral administration of AC induced apoptosis within the brain cells of rats by activating caspase-3. Co-administration of EG and AC has resulted in a reduced generation of caspase-3 as shown in table 2, and our result was also in agreement with the study carried out by Mesole et al. [14].
The results obtained from our study suggest that EG might exert its anti-apoptotic effects via the suppression of the caspase cascade activation.
The intrinsic pathway is coordinated by the Bcl-2 group of proteins. These group of proteins may be classified into two main sub-classifications based upon structural homology and these are anti and pro-apoptotic proteins. Maintaining equilibrium between anti/pro-apoptotic proteins is an essential factor which determines the fate of a cell. Negating the deteriorating effects of ROS within a cell is usually carried out by Bcl-2 within the cytosol by the reduction in the levels of various ROS [43]. The release of cytochrome c within the mitochondrial membrane is inhibited by Bcl-2. However, Bax on the other hand keeps in check the apoptotic process by the controlled release of cytochrome c into the cytosol as a result, leading to activation of caspase-3 [44]. Results obtained from this study shows that AC increased the levels of Bax and consequently lowered the levels of Bcl-2. Co-administration with EG significantly prevented the alteration in the levels of Bax and Bcl-2. Bax/Bcl-2 ratio shows the susceptibility of a cell to apoptosis. The higher the ratio the more susceptible a cell will be to apoptosis. Results obtained showed the group administered AC had a higher Bax/Bcl-2 ratio, while co-administration of EG and AC resulted in a significantly lower Bax/Bcl-2 ratio. Administration of EG only resulted in a Bax/Bcl-2 ratio, which was similar to the CTRL group. Result obtained was in agreement to the study carried out by Mesole et al. [14]. Administration of EG resulted to inhibition of toxicity as that resulted from AC administration. Scavenging nitrogen-based and oxygen reactants produced within the mitochondria is one major route by which EG stabilizes the mitochondrial membrane and as a result amplifying the anti-apoptotic signaling.
Extracts from the leaves of Moringa, Ocimum sanctum, cloves have also noted to be highly effective antioxidants by preventing the production of ROS and iNOS which are known to enhance the process of apoptosis in a cell [45]. Results obtained from our study are comparable to the studies which involved the plants mentioned above.
The first characterized neurotransmitter was acetylcholine with very high levels in the brain; acetylcholine is a very vital neurotransmitter for learning and memory [46]; that is the central cholinergic system that is vital for the regulation of cognitive functions [35]. Deficits in acetylcholine levels have been held responsible for senile dementia and other degenerative cognitive disorders, including Alzheimer’s disease [47].
The enzyme AChE is a vital biological element of the cell membrane contributing to its integrity. An increase in the levels of AChE activity results in a faster breakdown or degradation of acetylcholine and as a result a consequent diminished stimulation of acetylcholine receptors, which will produce a reduction of diverse cholinergic (learning and memory), and non-cholinergic (cell proliferation and neurite outgrowth) functions [48, 49].
In this study, oral administration of AC resulted in a significant increase in the level of AChE and this result was in agreement with the studies carried out by Singh et al. [50] who reported an increase in the level of AChE. Increase in the tissue activity of AChE will consequently result in faster degradation of acetylcholine. This faster breakdown of acetylcholine is very evident in the figure which revealed a reduced immunoreactivity using ACAT-1 immunohistochemical stain in the group administered 100 mg/kg of AC (Figure 7). Co-administration of EG and AC, however, resulted in an improved immunoreactivity which correlated to a reduced tissue level of AChE and within the group administered 300 mg/kg EG only revealed a high immunoreactivity which was comparable to the CTRL group.
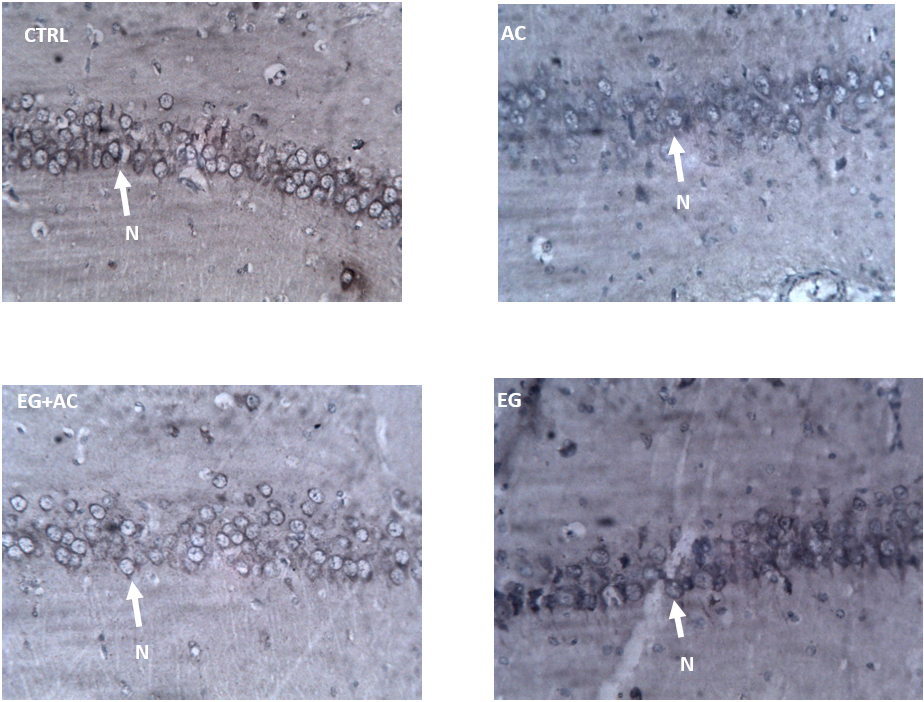
Figure 7: Effect of eugenol (EG) on the immunohistochemistry of the hippocampus (CA1) following the administration of aluminium chloride (AC).
Diminished levels of acetylcholine levels or increased AChE levels were found to be implicated in decreased cognitive function which was evident in the NOR. Result from NOR showed a significant reduction in time spent on exploring the novel object from day 7 to day 21 within the group administered AC. However, co-administration of EG and AC resulted in an increase in the time duration spent on exploring the novel object. An increase in time spent on a novel object is used as a yardstick for evaluating cognitive performance.
Author Contributions
Mesole SB: corresponding author and lead researcher, Ibegbu AO: supervision, Musa SA: supervision, Bauchi ZM: supervision, Agbon AN: data analysis and curation, Okpanachi OA: review and editing, Ivang A: research assistant.
Data Availability
Authors can confirm all relevant data are included in the article.
Conflicts of Interest
Authors declare no conflicts of interest.
References
- Inan-Eroglu E, Ayaz A. Is aluminum exposure a risk factor for neurological disorders? J Res Med Sci. 2018;23(1):51-65.
- Exley C. The pro-oxidant activity of aluminum. Free Radic Biol Med. 2004;36(3):380-87.
- Carson BL. Aluminum compounds. Review of toxicological literature, Abridged Final Report: p.84, 2000 integrated laboratory Systems, Research Triangle Park, North Carolina.
- Hirata-Koizumi M, Fujii, S, Ono, A, et al. Evaluation of the reproductive and developmental toxicity of aluminium ammonium sulfate in a two-generation study in rats. Food Chem Toxicol. 2011;(49)(9):1948-959.
- Mander P, Brown GC. Activation of microglial NADPH oxidase is synergistic with glial iNOS expression in inducing neuronal death: a dual-key mechanism of inflammatory neurodegeneration. J Neuroinflammation. 2005;2(1):1186-742.
- Suarez-Fernandez MB, Soldado AB, Sanz-Medel A, et al. Aluminum-induced degeneration of astrocytes occurs via apoptosis and results in neuronal death. Brain Res. 1999;835(2):125-36.
- Murakami K, Yoshino M. Aluminum decreases the glutathione regeneration by the inhibition of NADP-isocitrate dehydrogenase in mitochondria. J Cell Biochem. 2004;93(6):1267-271.
- Hussain A, Brahmbhatt K, Priyani A, et al. Eugenol enhances the chemotherapeutic potential of gemcitabine and induces anticarcinogenic and anti-inflammatory activity in human cervical cancer cells. Cancer Biother Radiopharm. 2011,;26(5):519-27.
- Institute of laboratory Animal Resources committee on care and use of laboratory animals (2011). Guide for the care and use of laboratory animals. National academies press Washington D.C.
- Al-Olayan EM, El-Khadragy MF, Moneim AE. The protective properties of melatonin against aluminium-induced neuronal injury. Int J Exp Pathol. 2015;96(3):196-202.
- Mæhre HK, Dalheim L, Edvinsen GK, et al. Protein determination—method matters. Foods. 2018;7(1):5.
- Okey SM, Ayo JO. The role of co-administration of ascorbic acid and zinc gluconate on brain biochemical changes in Wistar rats during the hot-humid season. Euro J Biotechnol Biosci. 2015;3(1):46-52.
- Kumar A, Khushboo, Pandey R, et al. Modulation of Superoxide Dismutase activity by Mercury, Lead, and Arsenic. Biol Trace Elem Res. 2020;196(2):654-61.
- Mesole SB, Alfred OO, Yusuf UA, et al. Apoptotic inducement of neuronal cells by Aluminium Chloride and the neuroprotective effect of eugenol in wistar rats. Oxid Med Cell Longev. 2020;8425643.
- Odinga T, Nwaokezi CO. Effect of Ricinodendron heudelotii seed extract on the oxidative stress biomarkers of diabetic albino rats. J Pharma Res Rev. 2020;4(19):1-6.
- Rui D, Yongjian Y. Aluminum chloride induced oxidative damage on cells derived from hippocampus and cortex of ICR mice. Brain Res. 2010;1324(1):96-102.
- El-Marasy SA, Abd-Elsalam RM, Ahmed-Farid OA. Ameliorative effect of Silymarin on Scopolamine-induced dementia in rats. Open Access Maced J Med Sci. 2018;6(7):1215-224.
- Zhao S, Wesseling S, Spenkelink B, et al. Physiologically based kinetic modelling based prediction of in vivo rat and human acetylcholinesterase (AChE) inhibition upon exposure to diazinon. Arch Toxicol. 2021;95(5):1573-593.
- Amirazodi F, Mehrabi A, Amirazodi M, et al. The combination effects of resveratrol and swimming HIIT exercise on novel object recognition and open-field tasks in aged rats. Exp Aging Res. 2020;46(4):336-58.
- de Almeida W, Confortim HD, Deniz BF, et al. Acrobatic exercise recovers object recognition memory impairment in hypoxic-ischemic rats. Int J Dev Neurosci. 2021;81(1):60-70.
- Labban S, Alghamdi BS, Alshehri FS, et al. Effects of melatonin and resveratrol on recognition memory and passive avoidance performance in a mouse model of Alzheimer’s disease. Behav Brain Res. 2021; 402:113100.
- Olajide OJ, Gbadamosi IT, Yawson EO. et al. Hippocampal degeneration and behavioral impairment during alzheimer-like pathogenesis involves glutamate excitotoxicity. J Mol Neurosci. 2021;71(6):1205-220.
- Lobine D, Sadeer N, Jugreet S, et al. Potential of medicinal plants as neuroprotective and therapeutic properties against amyloid-β-related toxicity, and glutamate-induced excitotoxicity in human neural cells. Curr Neuropharmacol. 2021: Online ahead of print.
- Jeon J, Bu F, Sun G, et al. Contribution of TRPC channels in neuronal excitotoxicity associated with neurodegenerative disease and ischemic stroke. Front Cell Dev Biol. 2021;8:618663.
- Ludolph AC, Riepe M, Ullrich K. Excitotoxicity, energy metabolism and neurodegeneration. J Inherit Metab Dis. 1993;16(4):716-23.
- Fulgenzi A, Vietti D, Ferrero ME. Aluminium involvement in neurotoxicity. Biomed Res Int. 2014;758323.
- Halliwell B, Gutteridge JMC. Free Radicals in Biology and Medicine. 3rd ed. New York: Oxford Science Publications; 2018.
- Norfaizatul SO, Zetty Akmal CZ, Noralisa AK, et al. Dual effects of plant antioxidants on neuron cell viability. J Medicinal Plants. 2010;9(6):113-23.
- Griñán-Ferré C, Bellver-Sanchis A, Izquierdo V, et al. The pleiotropic neuroprotective effects of resveratrol in cognitive decline and Alzheimer’s disease pathology: From antioxidant to epigenetic therapy. Ageing Res Rev. 2021;67:101271.
- Mesalam AA, El-Sheikh M, Joo MD, et al. Induction of oxidative stress and mitochondrial dysfunction by juglone affects the development of bovine oocytes. Int J Mol Sci. 2020;22(1):168.
- Yousef MI, El-Morsy AM, Hassan MS. Aluminium-induced deterioration in reproductive performance and seminal plasma biochemistry of male rabbits: protective role of ascorbic acid. Toxicology. 2005;215(1-2):97-107.
- Moneim AEA. Evaluating the potential role of pomegranate peel in aluminum-induced oxidative stress and histopathological alterations in brain of female rats. Biol Trace Elem Res. 2012;150(1):328-36.
- Romero A, Ramos E, de Los Rios C, et al. A review of metal-catalyzed molecular damage: protection by melatonin. J Pineal Res. 2014;56(4):343-70.
- Semiz, A., and Sen, A. Antioxidant and chemoprotective properties of Momordica charantia L. (Bitter melon) fruit extract. African J Biotechnol. 2007;6(3):273-77.
- Kankofer M. Superoxide dismutase and glutathione peroxidase activities in bovine placenta: spectrophotometric and electrophoretic analysis. Revue Méd Vét. 2002;153(2):121-24.
- Garabadu D, Shah A, Singh S, et al. Protective effect of eugenol against restraint stress-induced gastrointestinal dysfunction: Potential use in irritable bowel syndrome. Pharm Biol. 2015;53(7):968-74.
- Khan RA, Khan MR, Sahreen S. Brain antioxidant markers, cognitive performance and acetylcholinesterase activity of rats: efficiency of Sonchus asper. Behav Brain Funct. 2012;8(21):1-7.
- Yuan C-Y, Lee Y-J, Hsu G. Aluminum overload increases oxidative stress in four functional brain areas of Neonatal rats. J Biomed Sci. 2012;19(1):1-9.
- Mesole SB, Oseluka IA, Musa SA, et al. Histological and anti-oxidative effects of eugenol on aluminium chloride-induced neurotoxicity in Wistar rats. Nig J Neurosci. 2020;11(1):28-34.
- Adefegha SA, Oyeleye SI, Okeke BM, et al. Influence of eugenol on oxidative stress biomarkers in the liver of carrageenan-induced arthritis rats. J Basic Clin Physiol Pharmacol. 2018;30(2):185-93.
- Li J-M, Shah AM, Endothelial cell superoxide generation: regulation and relevance for cardiovascular pathophysiology. Am J Physiol Regul Integr Comp Physiol. 2004;287(5):R1014-30.
- Chiorcea-Paquim A-M, Oliveira-Brett AM. DNA electrochemical biosensors for In Situ probing of pharmaceutical drug oxidative DNA damage. Sensors. 2021;21(4):1125.
- Kumar V, Bal A, Gill KD. Impairment of mitochondrial energy metabolism in different regions of rat brain following chronic exposure to aluminium. Brain Res. 2008;1232(1):94-103.
- Akifusa S, Kamio N, Shimazaki Y, et al. Globular adiponectin-induced RAW 264 apoptosis is regulated by a reactive oxygen species-dependent pathway involving Bcl-2. Free Radic Biol Med. 2009;46(9):1308-316.
- Cai X, Zhang H, Tong D, et al. Corosolic acid triggers mitochondria and caspase-dependent apoptotic cell death in osteosarcoma MG-63 cells. Phytother Res. 2011;25(1):1354-361.
- Xu D-P, Li Y, Meng X, et al. Natural antioxidants in foods and medicinal plants: extraction, assessment and resources. Int J Mol Sci. 2017;18(1):96-128.
- Eu WZ, Chen Y-J, Chen W-T, et al. The effect of nerve growth factor on supporting spatial memory depends upon hippocampal cholinergic innervation. Transl Psychiatry. 2021;11(1):162.
- Kumar GP, Khanum F. Neuroprotective potential of phytochemicals. Pharmacogn Rev. 2012;6(12):81-90.
- Prasad SN, Muralidhara. Neuroprotective efficacy of eugenol and isoeugenol in Acrylamide-induced neuropathy in rats: behavioral and biochemical evidence. Neurochem Res. 2013;38(2):330-45.
- Singh V, Panwar R. In vivo antioxidative and neuroprotective effect of 4-allyl-2-methoxyphenol against chlorpyrifos-induced neurotoxicity in rat brain. Mol Cell Biochem. 2014;388(1):61-74.
